Impact Factor ISSN: 1449-2288
- Issue 11; 2026
- Issue 10; 2026
- Issue 9; 2026
- Issue 8; 2026
- Issue 7; 2026
- Volume 22; 2026
- Past Issues
- Advance Articles
- Editorial Board
- Cover Images
- Index & Coverage
- Cover Suggestion
- Special Issues
1. Introduction
2. The biology of HMGB1
3. HMGB1 and renal homeostasis
4. Pathogenic roles of HMGB1 in...
5. HMGB1 antagonists of...
6. Conclusions and perspectives
Funding
References

 Global reach, higher impact
Global reach, higher impactInt J Biol Sci 2023; 19(15):5020-5035. doi:10.7150/ijbs.87964 This issue Cite
Review
Targeting HMGB1: A Potential Therapeutic Strategy for Chronic Kidney Disease
Tongtong Liu1#, Qian Li1#, Qi Jin1, Liping Yang1, Huimin Mao1, Peng Qu2, Jing Guo3, Bo Zhang2, Fang Ma1, Yuyang Wang1, Liang Peng2 ![]() , Ping Li2
, Ping Li2 ![]() , Yongli Zhan1
, Yongli Zhan1 ![]()
1. Guang'anmen Hospital, China Academy of Chinese Medical Sciences, Beijing, China.
2. China-Japan Friendship Hospital, Institute of Medical Science, Beijing, China.
3. Institute of Basic Research in Clinical Medicine, China Academy of Chinese Medical Sciences, Beijing, China.
# co-author: Tongtong LIU and Qian LI contributed equally to the article.
Received 2023-7-10; Accepted 2023-9-18; Published 2023-9-25
Abstract

High-mobility group protein box 1 (HMGB1) is a member of a highly conserved high-mobility group protein present in all cell types. HMGB1 plays multiple roles both inside and outside the cell, depending on its subcellular localization, context, and post-translational modifications. HMGB1 is also associated with the progression of various diseases. Particularly, HMGB1 plays a critical role in CKD progression and prognosis. HMGB1 participates in multiple key events in CKD progression by activating downstream signals, including renal inflammation, the onset of persistent fibrosis, renal aging, AKI-to-CKD transition, and important cardiovascular complications. More importantly, HMGB1 plays a distinct role in the chronic pathophysiology of kidney disease, which differs from that in acute lesions. This review describes the regulatory role of HMGB1 in renal homeostasis and summarizes how HMGB1 affects CKD progression and prognosis. Finally, some promising therapeutic strategies for the targeted inhibition of HMGB1 in improving CKD are summarized. Although the application of HMGB1 as a therapeutic target in CKD faces some challenges, a more in-depth understanding of the intracellular and extracellular regulatory mechanisms of HMGB1 that underly the occurrence and progression of CKD might render HMGB1 an attractive therapeutic target for CKD.
Keywords: HMGB1, CKD, renal homeostasis, AKI-to-CKD transition, therapeutic strategy
1. Introduction
Chronic kidney disease (CKD), a devastating disease affecting human health worldwide, is characterized by progressive and irreversible nephron loss, reduced renal regenerative capacity, microvascular damage, changes in inflammation, metabolic and oxidative stress, and fibrosis, ultimately leading to renal failure and end-stage renal disease (ESRD)[1, 2]. CKD affects approximately 10-14% of the global population and is the leading cause of ESRD and premature death[3]. The contribution of CKD to global mortality is rapidly increasing due to the rising prevalence of diabetes, hypertension, obesity, and an aging population[4]. However, current treatments have limited efficacy and merely delay disease progression. Therefore, it is essential to identify new potential therapeutic targets to halt or reverse CKD progression.
High-mobility group protein box 1 (HMGB1) is a member of the high-mobility group proteins with secretory and intracellular activities[5]. HMGB1 is ubiquitously expressed in almost all cell types and is involved in cellular damage and repair. The biological activity of HMGB1 depends on its subcellular localization, context, and post-translational modifications (PTMs). HMGB1 acts as a DNA chaperone in the nucleus and is involved in DNA repair, chromatin remodeling, nucleosome assembly, and telomere maintenance. In the cytoplasm, HMGB1 acts as an autophagy maintainer and mitochondrial homeostasis regulator to regulate cell death. Extracellularly, HMGB1 acts as a damage-associated molecular pattern (DAMPs) or alarmin to activate the immune response and promote cell migration and proliferation[6]. HMGB1 has been discovered for 50 years (Figure 1). In recent years, an increasing number of HMGB1 inhibitors have shown promising therapeutic potential for a variety of diseases[7, 8]. HMGB1 plays an indispensable role in the pathogenesis and progression of CKD. The kidney is the best responder to HMGB1 because of the largest changes in HMGB1 in kidney tissue in the early stage of hemorrhagic shock[9]. Under CKD conditions, HMGB1 is elevated in the plasma, serum, and urine and is closely related to the progression and prognosis of CKD. As a core player, activated HMGB1 participates in multiple key events of CKD progression through the activation of downstream signals, including renal inflammation, the development of persistent fibrosis, renal aging, AKI-CKD transition, and important cardiovascular complications. Interestingly, studies have shown that HMGB1 deletion in renal tubules has no noticeable effect on renal injury in the early stage after unilateral ureteral obstruction (UUO) but greatly alleviates renal interstitial fibrosis in the late/subacute stage[10], suggesting that HMGB1 may play additional roles in CKD than differ from those in acute kidney injury.
Timeline of landmark achievements of HMGB1 in CKD in the past 50 years.

In this review, we evaluate the links between HMGB1 and CKD, starting with a description of the biological characteristics of HMGB1 in CKD and its regulatory role in renal homeostasis, followed by valuable preclinical and clinical evidence, summarizing how HMGB1 regulates key events related to CKD to affect its progression and prognosis. Finally, we describe strategies to reduce or inhibit HMGB1 in CKD, providing insights into the innovation of therapeutic strategies targeting HMGB1 in CKD.
2. The biology of HMGB1
The HMGB protein family is the most abundant among high-mobility groups. Four members (HMGB1, HMGB2, HMGB3 and HMGB4) of the mammalian HMGB family have been identified so far; among them, HMGB1 shows the highest expression. HMGB1, a non-histone nuclear protein, was first discovered in 1973 and named for its high electrophoretic mobility. HMGB1 is highly evolutionarily conserved, as evidenced by the 99% homology between rodent and human amino acid sequences. HMGB1 is essential for life as mice with systemic HMGB1 deletions die from hypoglycemia shortly after birth[11]. Of note, HMGB1 can cross organelles from the nucleus at higher concentrations into the cytoplasm in response to stress injury within 1-2 seconds[12].
2.1 The structure and distribution of HMGB1
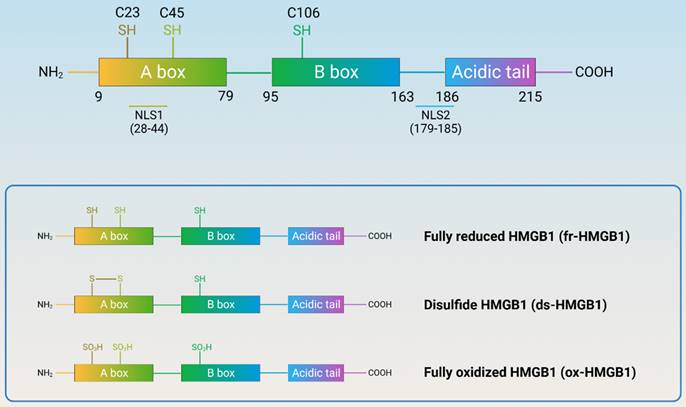
Human HMGB1 consists of 215 amino acid residues that form two homologous DNA-binding domains (A-box and B-box), a negatively charged C-terminal acidic tail, and a short but functionally significant N-terminal region (Figure 2). HMGB1 contains three redox-sensitive cysteine residues (C23, C45, and C106). C23 and C45 can form intramolecular disulfide bonds, while C106 is unpaired. Based on the redox status of the three cysteine residues, HMGB1 can be classified into three subtypes: fully reduced HMGB1 (fr-HMGB1, with three conserved cysteine residues containing thiol groups), disulfide HMGB1 (ds-HMGB1, partially oxidized), and fully oxidized HMGB1 (ox-HMGB1, sulfonyl HMGB1). fr-HMGB1 can bind to other chemokines to promote immune cell migration and tissue regeneration. ds-HMGB1 can activate immune cells to produce cytokines/chemokines and exhibit a higher affinity for the nuclear export of CRM1[13]. However, ox-HMGB1 exhibited no chemokine or cytokine activity. Importantly, the exchange between fr-HMGB1 and ds-HMGB1 is reversible, while that with ox-HMGB1 is irreversible[14].
Structure and redox reaction of HMGB1. (HMGB1 is composed of A-box, B-box, C-terminal acidic tail, and a short but functionally significant N-terminal region, with nuclear localization signals and three redox-sensitive cysteine residues. HMGB1 can be classified into three subtypes: fully reduced HMGB1, disulfide HMGB1, and fully oxidized HMGB1.)
2.2. The distribution and function of HMGB1
HMGB1 is highly expressed in various kidney cells, and its role in CKD pathogenesis depends on its subcellular localization (Figure 3). In the nucleus, HMGB1 promotes the repair of damaged DNA and the maintenance of nucleosome homeostasis and telomere homeostasis. In particular, the retention of HMGB1 in the nucleus improves the differentiation of peripheral B cells and the phagocytic capacity and chemotactic response of macrophages[15, 16]. In the cytoplasm, HMGB1 is primarily involved in regulating autophagy, mitochondrial function, and apoptosis. Extracellular HMGB1 primarily serves as a DAMP and participates in many immune responses by promoting immune cell maturation, activation, and cytokine production[17]. More importantly, extracellular HMGB1 is associated with cell death. extracellular HMGB1 can be internalized and targeted to lysosomes, inducing lysosomal membrane permeabilization (LMP) and accelerating subsequent cell death[18, 19].
The distribution and function of HMGB1. (HMGB1 can cross organelles from the nucleus at higher concentrations into the cytoplasm in response to stress injury. The function of HMGB1 is related to its subcellular structure. In the nucleus, HMGB1 plays an important role in DNA replication and repair, chromatin remodeling, nucleosome assembly, and telomere maintenance; In the cytoplasm, HMGB1 is primarily involved in regulating autophagy, mitochondrial function, and apoptosis; Extracellular HMGB1 primarily serves as a DAMP and participates in many immune responses, can also promote cell migration and proliferation.)
2.3. The modification and regulation of HMGB1
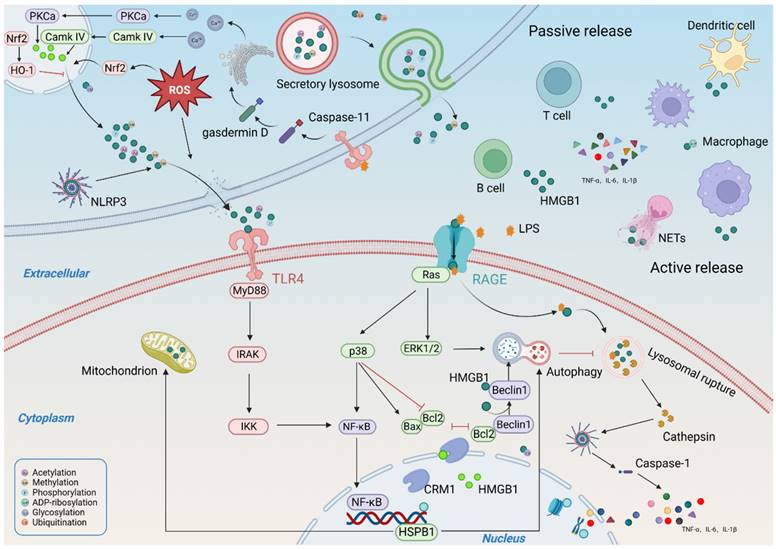
The localization and activity of HMGB1 are affected by PTMs[20], including acetylation, methylation, phosphorylation, poly-ADP-ribosylation, and glycosylation. Acetylation enhances the ability of HMGB1 to bend DNA and prevents HMGB1 from re-entering the nucleus[21, 22]. Resveratrol (a natural SIRT1 agonist) pretreatment promoted the nuclear retention of HMGB1 by reducing HMGB1 acetylation, thereby improving renal inflammation and tubular injury[23]. Methylation changes the conformation of HMGB1 and weakens its DNA-binding activity, allowing its massive passive diffusion into the cytoplasm and subsequent secretion extracellularly[24]. Phosphorylation also limits the nuclear localization of HMGB1 by modifying its two NLS [25]. Poly (ADP)-ribose polymerase (PARP) also promotes the nuclear release of HMGB1 into the extracellular environment[26]. Poly-(ADP)-ribosylated HMGB1 not only downregulates gene transcription[27, 28] but also inhibits efferocytosis in macrophages to a significant extent, thereby promoting inflammation[29]. In turn, the deletion of HMGB1 leads to excessive PARP-1 activation, which exacerbates mitochondrial damage and cell death[30]. In addition, PARP-1 also induced the release of HMGB1 from proximal tubular cells[31]. Glycosylation plays a crucial role in HMGB1secretion. N-glycosylation weakens the binding of HMGB1 to DNA and enhances its binding to the nuclear export protein CRM1, a prerequisite for HMGB1 cytoplasmic transport and extracellular secretion[32]. In addition, N-glycosylation of HMGB1 leads to reduced binding to glycyrrhizin, an HMGB1 inhibitor[33]. Recent studies have demonstrated that O-glcnacylation can also modify HMGB1, resulting in its reduced ability to repair DNA[34]. Recently, S-nitrosylation has been shown to promote HMGB1 secretion and proinflammatory effects[35]. Ubiquitination modification helps promote HMGB1 degradation and improves disease progression[36]. Ubiquitin-specific protease-12 deubiquitinates and stabilizes HMGB1 to promote autophagy by interacting with HMGB1[37]. However, another study showed that compared with the ubiquitination pathway, the autophagy-lysosome pathway plays a major role in HMGB1 degradation, and the activation of autophagy and an increase in CTSB promote HMGB1 degradation and nuclear translocation[38].
2.4. Secretion and release of HMGB1
Under the action of various stressors (such as hypoxia, cytokines, chemokines, and uremic toxins) in CKD, HMGB1 is secreted externally through an unconventional protein secretion pathway in an active or passive manner, rather than through the conventional endoplasmic reticulum-Golgi pathway[6]. At present, two main secretion pathways of HMGB1 have been proposed. One is the direct, pore-mediated secretion of HMGB1 by pyroptosis or activated target cells[39]; the other is secretory autophagy, which packages HMGB1 into intracellular vesicles (such as lysosomes or autophagosomes) and releases HMGB1 through exocytosis[40]. However, these two pathways are difficult to distinguish because they occur simultaneously in most pyroptotic cells[39]. The regulatory mechanism underlying HMGB1 secretion is complex and involves several pathways. Oxidative stress is known to be an important factor in regulating HMGB1 secretion[41]. HMGB1 is secreted through a ROS-dependent mechanism under hypoxia, and targeted inhibition of ROS production significantly reduces HMGB1 secretion. Nuclear factor erythroid 2-related factor 2 (Nrf2) is a key transcription factor regulated by oxidative stress. Nrf2 knockdown abolishes the regulatory effect of antioxidants on HMGB1[42]. Notably, the regulation of HMGB1 secretion by oxidative stress is driven by calcium signaling[43]. The inhibition of calcium/calmodulin-dependent kinase resulted in a significant reduction in HMGB1 secretion. Calcium overload promotes the release of phosphorylated HMGB1[44]. Moreover, HMGB1 release mediates calcium influx by promoting calcium channel activation [45]. In addition, the cytoplasmic translocation and secretion of HMGB1 are also tightly regulated by the nuclear export protein CRM1. The inhibition of CRM1 expression significantly reduced circulating HMGB1 levels[46]. In addition, several cathepsin family members also promote HMGB1 secretion. Under stress, the permeability of the lysosomal membrane is altered, leading to the release of cathepsins and other hydrolases in the cytoplasm and their subsequent translocation to the nucleus, inducing the formation of the NLRP3 inflammasome complex, ultimately leading to increased HMGB1 secretion[47, 48]. The released HMGB1 targets LPS internalization into lysosomes through RAGE and mediates lysosomal leakage, which activates caspase 11 and promotes pyroptosis[18]. Targeted inhibition of HMGB1 binding to LPS improves lysosomal rupture and attenuates caspase 11-mediated sepsis-related lethality[49]. A recent study showed that TLR4 also increases the expression of caspase 11 through LPS uptake and that activated caspase 11 promotes the cleavage of gasdermin D, resulting in increased calcium release from the endoplasmic reticulum, which in turn promotes HMGB1 secretion[50].
3. HMGB1 and renal homeostasis
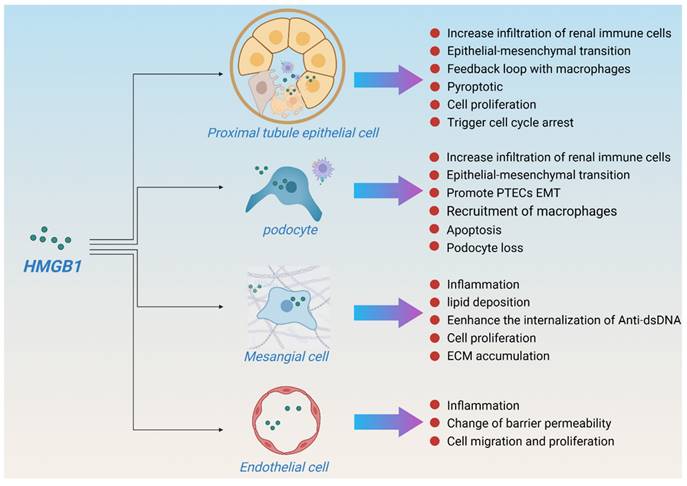
HMGB1 is expressed in a variety of kidney cell types, including glomerular epithelial cells (podocytes), endothelial cells, tubular cells, inflammatory mononuclear phagocytes, and lymphocytes. In case of injury, renal tubular epithelial cells and podocytes are the main sources of HMGB1, and mesangial and endothelial cells also express HMGB1[51], which in turn promotes apoptosis and renal inflammation. Although macrophage-derived HMGB1 plays an important role in many diseases[52, 53], studies have shown that macrophage-derived HMGB1 does not aggravate renal fibrosis after UUO[54]. In contrast, the deletion of bone marrow-derived RAGE contributed to the improvement of renal function in a DKD mouse model[55], indicating that macrophages may only be effectors of HMGB1 rather than the main secretory source during kidney injury, especially in CKD. In this section, we summarize the important regulatory effects of HMGB1 on various intrinsic kidney cell types to maintain kidney homeostasis. The effects of HMGB1 on intrinsic renal cells in various kidney disease models are summarized in Table 1 and Figure 4.
HMGB1 effects in renal resident cells.
| Cell types | Types of study | Experimental models | Pathway | HMGB1-mediated effects | References |
|---|---|---|---|---|---|
| Proximal tubule epithelial cell | In vitro, In vivo | UUO | TNFα/Casp3/GSDME/HMGB1 | Promote inflammation, PTEC damage and fibrosis | [10] |
| In vitro, In vivo | CsA-induced renal injury | HMGB1/TLR4 | Promote inflammation and fibrosis | [63] | |
| In vivo | HG-induced HK-2 | HMGB1/TLR4/Syk | Promote NF-κB activation and TGF-β1 production | [171] | |
| In vitro, In vivo | Ang II-induced renal injury | NLRP3/HMGB1 | Promote EMT and fibrosis | [172] | |
| In vitro, In vivo | FLCs-induced renal injury | STAT1/HMGB1/TLR | Promote inflammation and PTEC damage | [58] | |
| In vivo | AGE-induced HK-2 | HMGB1/RAGE | Promote the expression of CTGF and TGF-β | [59] | |
| In vivo | HMGB1-induced HK-2 | HMGB1/RAGE | Promote EMT | [60] | |
| In vitro, In vivo | UUO | C3 / HMGB1 / TGF-β1 | Promote EMT and fibrosis | [61] | |
| In vitro, In vivo | STZ-induced DKD | HMGB1/TLR2/4/NF-κB | Promote inflammation | [161] | |
| In vitro, In vivo | HFD-fed OLETF rats | NLRP3/HMGB1 | Promote inflammation and PTEC damage | [173] | |
| In vitro, In vivo | IMI-induced renal injury | HMGB1-RAGE/TLR4-NF-κB | Promote PTEC Ferroptosis and pyroptosis | [62] | |
| In vivo | AAs-induced HK-2 | ROS/HMGB1/mt DNA/ TLRs | EMT and mitochondrial dysfunction | [64] | |
| In vitro, In vivo | CNIs-induced renal injury | / | Promote PTEC mitochondrial dysfunction and bioenergetic reprograming | [65] | |
| In vivo | CaCl2-induced HK-2 | HMGB1/TLR4/NF-κB | Promote inflammation and autophagy | [143] | |
| In vitro, In vivo | UUO | / | Promote fibrosis | [115] | |
| Podocyte | In vitro, In vivo | db/db mice | / | Promote podocyte apoptosis and EMT | [71]. |
| In vitro, In vivo | ADR-induced renal injury | / | Promote podocyte injury and proteinuria | [72] | |
| Mesangial cell | In vitro, In vivo | MRL/lpr mice | TLR2/MyD88/NF-κB | Promote glomerular mesangial matrix deposition | [76] |
| In vivo | HG-induced SV40 MES 13 | HMGB1/NF-κB | Promote inflammation | [174] | |
| In vivo | HG-induced SV40 MES 13 | HMGB1/TLR4/NF-κB | Promote proliferation, oxidative stress, ECM accumulation, and inflammation in mesangial cells | [175] | |
| In vivo | HMGB1-induced SV40 MES 13 | HMGB1/PTEN/PI3K/Akt | Promote proliferation in mesangial cells | [176] | |
| In vitro, In vivo | db/db mice | Hspa9/HMGB1 | Promote proliferation and fibrosis in mesangial cells | [177] | |
| In vivo | IFN-γ- induced MMC | JAK2 / STAT1 | Promote lipogenesis in mesangial cells | [74] | |
| In vivo | TWEAK and anti-dsDNA IgG-induced MMC | TWEAK/Fn14; NF‐κB/PI3K/AKT | Promote anti-dsDNA IgG penetration into mesangial cells | [75] | |
| In vitro, In vivo | MRL/lpr mice | TLR2/4 and RAGE | Promote inflammation | [77] | |
| In vitro, In vivo | NAFLD + BDCM-induced renal injury | HMGB1/TLR4 | Promote mesangial cell activation | [78] | |
| In vivo | HMGB1-induced MMC | PI3K/Akt | Promote proliferation of mesangial cell | [79] | |
| In vivo | HG-induced SV40 MES 13 | TLR4/NF-κB | Promote ferroptosis in mesangial cells | [80] | |
| Endothelial cell | In vivo | HMGB1-induced HUVECs | / | Promote angiogenesis in Endothelial Cells | [84] |
| In vitro, In vivo | MRL/lpr mice | TLR4/MyD88 | Induced glomerular endothelial cell injury | [86]. | |
| In vivo | sera from AAV patients GEnCs | HMGB1/TLR4 | Induced glomerular endothelial cell injury | [87] |
HG: high glucose; STZ: Streptozocin; FLCs: free light chains; EMT: epithelial-mesenchymal transition; IMI: imidacloprid; CsA: Cyclosporine A; (mt DNA: mitochondrial DNA; AAs: aristolochic acids; HFD: High fat diet; ADR: adriamycin.
HMGB1 and renal homeostasis. (HMGB1 is expressed in a variety of kidney cell types, especially in proximal tubule epithelial cell and podocyte. In case of injury, renal tubular epithelial cells and podocytes are the main sources of HMGB1, and mesangial and endothelial cells also express HMGB1. HMGB1 mediates kidney damage and repair through multiple pathways to maintain renal homeostasis.)
3.1 HMGB1 and proximal tubule epithelial cell
Proximal tubular epithelial cells (PTECs) are the major epithelial cell type in the cortex. The effects of HMGB1 on PTECs function have been extensively studied. HMGB1-mediated tubular injury and renal fibrosis are seemingly hallmarks of chronic processes[10]. In the early stages of PTEC injury, HMGB1 secretion promotes rapid γδ T-cell infiltration and mediates an early immune response to renal injury[56]. In the late stage of kidney injury, PTECs reduce HMGB1 secretion through locally produced propertin (a positive regulator of the alternative complement pathway), thereby reducing macrophage infiltration and enhancing the phagocytic capacity of PTECs, which in turn curtails apoptosis and kidney inflammation[57]. In addition, free light chains (FLCs) promote the secretion of HMGB1 by PTECs and the expression of TLR2, TLR4, and TLR6, resulting in an overload of the endocytic pathway of FLCs, which triggers inflammation and cell damage[58]. HMGB1 also promotes transforming growth factor β (TGF-β) and connective tissue growth factor (CTGF) expression and induces epithelial-mesenchymal transition (EMT) of PTECs, ultimately accelerating renal fibrosis[59-61].
Notably, HMGB1 is involved in renal tubular injury caused by various nephrotoxic drugs. For instance, imidacloprid stress induces Nrf2 inactivation and mediates HMGB1/RAGE/TLR4 signaling activation, thereby triggering iron death and leading to the initial wave of death that fuels pyroptosis and exacerbates renal dysfunction[62]. Similarly, cyclosporine, aristolochic acid I, and calcineurin inhibitors also promote HMGB1 secretion by tubular cells, aggravating tubular injury and renal fibrosis[63-65], suggesting that HMGB1 might serve as an early indicator and marker of progressive nephrotoxicity.
Although neutralization of extracellular HMGB1 is beneficial, intracellular HMGB1 seems to play an additional role in renal tubular injury[51]. The induction of increased HMGB1 levels in the plasma and urine by remote ischemic preconditioning, but not by increased infiltration of renal immune cells, reportedly reduced the risk of renal injury[66]. Indeed, recent studies have also found that HMGB1 plays a dual role in renal tubules. It has been found that HMGB1 binds to TLR4 on PTECs to trigger transient protective G1 cell cycle arrest, providing renal protection[67], while TLR4 activation on non-renal cells has been shown to contribute to renal injury[66].
3.2 HMGB1 and podocyte
Podocytes are terminally differentiated glomerular epithelial cells that play a key role in maintaining the glomerular filtration barrier[68]. Podocytes are non-professional antigen-presenting cells that are both the target of inflammatory injury and active participants[69]. Damaged podocytes are one of the primary sources of renal HMGB1 secretion. Under injury conditions, podocytes promote renal injury by secreting HMGB1 to promote the EMT of PTECs, mitochondrial damage, and apoptosis[70]. Targeting HMGB1 inhibition or depletion ameliorates podocyte injury and EMT by regulating autophagy homeostasis[71]. CLEC14a is a single-pass transmembrane glycoprotein that exerts a protective effect on podocytes. CLEC14a ameliorates podocyte injury by improving NF-κB signaling and early growth response protein 1 signaling via directly binding to HMGB1 and inhibiting its release[72]. In addition, deleting bone marrow-derived RAGE improved podocyte loss following streptozocin (STZ) induction[55].
3.3 HMGB1 and mesangial cell
Mesangial cells (MCs) play an important role in maintaining the structural integrity of the glomerular microvascular bed and mesangial matrix homeostasis[73]. HMGB1 is an important mediator of MC activation. HMGB1 mediates lipid deposition in MCs by promoting the transcription and expression of sterol regulatory element-binding protein-1 and fatty acid synthase [74]. In lupus nephritis (LN), HMGB1 also enhances the internalization of anti‐double‐stranded DNA (dsDNA) IgG in MCs by binding to dsDNA IgG and activates the MyD88/NF-κB pathway[75, 76], which exhibits a synergistic proinflammatory effect that mediates the activation of MCs[77], leading to renal tubular cell death and increased cytokine release[78], thereby aggravating proteinuria, glomerulosclerosis, and renal fibrosis in LN[76, 79]. The depletion of HMGB1 in MCs inhibits iron death and improves MC proliferation by regulating Nrf2 signaling[80].
3.4 HMGB1 and endothelial cell
The population of ECs in the kidney is remarkably diverse, and approximately 24 morphologically and functionally heterogeneous EC types have been identified[81]. ECs are extremely sensitive to stress, and disruption of endothelial function is considered an early event in kidney injury[82]. At the early stage of kidney injury, HMGB1 released first exacerbates kidney injury by interacting with TLR4 in renal ECs (much earlier than in renal tubular epithelial cells), leading to EC activation and upregulating the expression of adhesion molecules[83]. HMGB1 is an important regulator of ECs. On the one hand, HMGB1 induces a proinflammatory response in ECs, leading to early changes in barrier permeability in ECs. On the other hand, the internalization of HMGB1 into ECs promotes the expression of vascular endothelial growth factor, which in turn promotes EC migration and proliferation[84, 85]. HMGB1 is highly expressed in the glomerular ECs of patients with LN. HMGB1 promotes the permeability of ECs and the shedding of the glycocalyx in the glomerulus and disrupts intercellular tight junctions and cytoskeleton arrangement, thus aggravating LN-related proteinuria[86]. Similarly, HMGB1 mediates myeloperoxidase (MPO)-antineutrophil cytoplasmic antibody (ANCA)-induced EC activation and glomerular damage by triggering moesin phosphorylation and secretion and promoting cross-reactivity between moesin and the anti-MPO antibody[87].
4. Pathogenic roles of HMGB1 in CKD
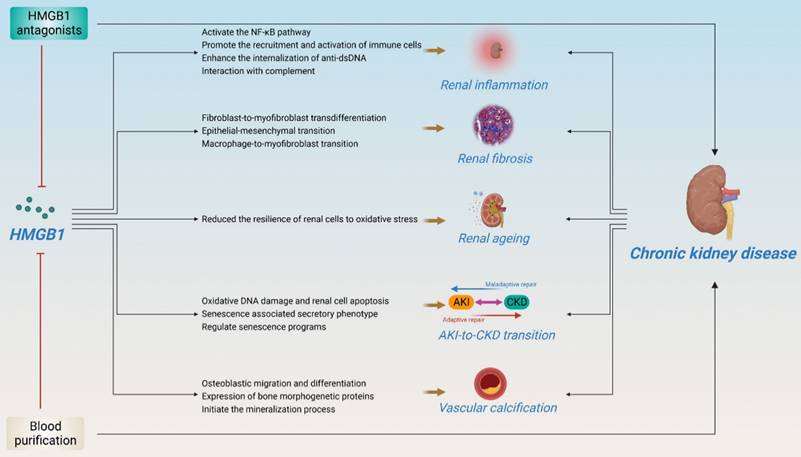
HMGB1 is involved in kidney disease progression. Although the important role of HMGB1 in kidney disease has been elegantly described in several reviews[88, 89], recent research advances suggest that HMGB1 plays an important role in kidney disease, especially in CKD, including kidney inflammation, fibrosis, ageing, AKI-to-CKD transition, vascular calcification, and renal replacement therapy, anticipating that strategies to block the interaction between HMGB1 and its receptor may be effective in preventing the development of CKD (Figure 5).
Pathogenic roles of HMGB1 in CKD. (HMGB1 plays an important role in kidney disease, especially in CKD, including kidney inflammation, fibrosis, ageing, AKI-to-CKD transition, vascular calcification, and renal replacement therapy. Several HMGB1 inhibitors and hemodialysis have shown potential therapeutic effects in improving CKD.)
4.1 The clinical value of HMGB1 in CKD
HMGB1 has been confirmed to be associated with the occurrence, progression, and prognosis of CKD in multiple clinical studies (Table 2). A study including 177 CKD patients found that HMGB1 was significantly elevated in patients with CKD and correlated with estimated glomerular filtration rate (eGFR) and markers of inflammation and malnutrition[90, 91]. An observational study including 20 patients with non-diabetic nephropathy found that serum HMGB1 was significantly elevated in CKD and independently correlated with the accumulation of asymmetric dimethylarginine, indicating that HMGB1 is actively involved in CKD progression and might lead to the development and progression of cardiovascular diseases (CVDs)[92]. In terms of pathological typing, a study including 258 patients with chronic glomerulonephritis (GN) found that HMGB1 tended to be significantly elevated in the serum of patients with ANCA-GN, Henoch-Schonlein purpura nephritis, and IgA nephropathy with glomerular crescents[93].
Clinical studies of HMGB1 in CKD
| CKD population | sample size (CKD/HC) | Measurement method of HMGB1 | Sample source | Major findings | References |
|---|---|---|---|---|---|
| GN | 258/49 | ELISA | Serum | HMGB1 was expressed in the sera of patients with renal diseases who underwent renal biopsies, especially among those who had vasculitis including ANCA-GN, Henoch-Schonlein purpura nephritis, and IgAN with glomerular crescents. | [93] |
| CKD | 177/48 | ELISA | Serum | HMGB-1 is elevated significantly in CKD patients and correlates with GFR as well as markers of inflammation and malnutrition. | [90] |
| CKD | 289/61 | ELISA | Serum | HMGB1 levels were significantly higher in CKD patients and related to disease stage | [91] |
| CKD | 20/20 | ELISA | Serum | HMGB-1 were independently associated with asymmetric dimethylarginine. | [92] |
| LN | 50(SLE)/50 | ELISA | Serum | Patients with LN had significantly higher serum HMGB1, and correlated positively to the SLE Disease Activity Index. | [94] |
| LN | 70(SLE)/35 | WB and ELISA | Serum | Serum HMGB1 levels are related to SLEDAI scores and proteinuria. | [95] |
| LN | 69(SLE)/17 | WB | Serum And Urine | Serum and urinary levels of HMGB1 were significantly increased in patients with active LN. | [96] |
| LN | 61(SLE)/14 | WB | Urine | HMGB1 is elevated in the urine of patients with active LN, and associated with LN class. | [97] |
| LN | 44(LN)/16(SLE) | Flow cytometry | Pbmcs And Urine | High frequencies of MP-HMGB1 in urine of LN patients | [98] |
| LN | 35(LN)/0 | WB | Serum | serum levels of HMGB1 were increased in LN, and there was no change after immunosuppressive therapy. | [99] |
| ANCA | 74(active AAV)/65(active AAV) | ELISA | Plasma | plasma levels of HMGB1 correlated with initial serum creatinine, and estimated glomerular filtration rate. | [100] |
| ANCA | 25/13 | WB | Serum | HMGB1 is significantly increased in AAV with renal involvement. | [101] |
| ANCA | 51(VAs)/46(HC) | ELISA | Serum | positive correlation between serum HMGB1 levels and Scr, and 24-hour proteinuria | [102] |
| CAPD | 62/31 | ELISA | Serum | HMGB-1 was elevated significantly in CAPD patients and correlated with indicators of inflammation and malnutrition. | [136] |
| ESRD | 151(HD)/ 102(PD) | ELISA | Serum | Serum level of HMGB1 in patients on HD was higher than PD, and patients with higher HMGB1 had more complications than patients with lower HMGB1, but there was no difference for the survival rate. | [138] |
GN: glomerulonephritis; CKD: chronic kidney disease; LN: lupus nephritis; ANCA: antineutrophil cytoplasmic antibody; CAPD: continuous ambulatory peritoneal dialysis; ESRD: end-stage renal disease; WB: western blot; ELISA: enzyme linked immunosorbent assay
In addition, HMGB1 shows good discriminative ability for LN. Serum HMGB1 levels are significantly elevated in patients with LN and correlate with SLE activity[94, 95]. Serum HMGB1 levels also positively correlate with proteinuria in patients with LN [94, 95]. However, in another study involving 69 patients with SLE, no significant correlation was found between serum HMGB1 and proteinuria [96]. In the urine of patients with LN, HMGB1 was also significantly elevated and correlated with the LN class, with higher levels of urinary HMGB1 in patients with LN class V[97]. Another study found that microparticle (MPs)-HMGB1 was elevated in the circulation and urine of patients with LN, and MP-HMGB1 in urine showed good discriminative ability for the presence of LN and disease activity[98]. Notably, at the end of follow-up, immunosuppressive treatment only reduced HMGB1 expression in the serum and renal tissues of class IV LN patients, whereas HMGB1 levels in other patients with LN did not change significantly before and after treatment, possibly due to persistent chronic inflammation[99].
The correlation between HMGB1 and renal injury in patients with ANCA-associated vasculitis (AAV) is particularly strong. A study that collected plasma samples from 74 patients with active AAV and 65 patients with remission AAV found that circulating HMGB1 levels were associated with renal involvement and that plasma HMGB1 levels significantly correlated with initial serum creatinine and eGFR[100]. Similarly, in a study that included 30 patients with AAV, HMGB1 was significantly elevated in AAV patients with renal involvement and continued to increase with disease activity[101]. In a study including 51 patients with AAV, serum HMGB1 was found to correlate significantly with disease activity and renal involvement and positively correlated with serum creatinine and 24-hour urinary protein levels in patients with AAV[102]. Similarly, urine HMGB1 levels also are associated with renal involvement in patients with AAV [103].
4.2 Renal inflammation and fibrosis
Patients with CKD experience persistent inflammation in the early stages, which determines the progression of most kidney diseases. Cells exposed to the kidney disease environment undergo phenotypic changes and overproduce proinflammatory cytokines, which in turn contribute to the recruitment of cells involved in innate and adaptive immune responses, further amplifying inflammation and damage to the kidney[104]. HMGB1 is an emerging mediator of renal inflammation. Mechanistically, On the one hand, HMGB1 activates the NF-κB pathway by interacting with RAGE and TLR4 in kidney cells[105, 106]; On the other hand, by promoting the recruitment and activation of immune cells, including macrophages[107], dendritic cells and B cells[108]. In LN, the released HMGB1 also contributes to the endocytosis of extracellular accumulated DNA and the activation of cyclic GMP-AMP synthase signaling pathway, and the subsequent secretion of IFN-I, leading to the expansion of downstream inflammation[109]. In addition, HMGB1 also amplifies renal inflammation through the interaction with complement. In ANCA, HMGB1 promoted C5a-mediated translocation of ANCA antigens and neutrophil activation, thereby aggravating renal involvement[110]. Importantly, HMGB1 mediated inflammation is an important factor driving renal fibrosis[111]. The severity of renal fibrosis is positively correlated with the activation of HMGB1/TLR2/TLR4 signaling[112]. Specifically, HMGB1 promotes the expression of TGF-β and CTGF by activating multiple inflammatory pathways[113, 114], which in turn promotes fibroblast-to-myofibroblast transdifferentiation and EMT[115, 116], accelerating renal fibrosis. In addition, HMGB1 promote the recruitment and activation of macrophages in the early stages of UUO and induce macrophage-to-myofibroblast transition, thereby promoting renal fibrosis[117]. Interestingly, surfactant protein A, a novel protein factor, can block TGF-β1 expression and renal fibroblast transdifferentiation by binding HMGB1, thereby improving renal fibrosis[115]. Therefore, targeted inhibition of HMGB1 may be a good strategy for improving renal fibrosis.
4.3 AKI-to-CKD transition
AKI leads to a significantly higher risk of CKD and ESRD, as well as higher mortality[118]. Therefore, preventing the transition from AKI to CKD is essential. Maladaptive repair and increased irreversible renal fibrosis after AKI are the main causes of CKD[119], including tubular epithelial cell injury, endothelial dysfunction, microvascular rarefaction, and inflammatory progression. Recent evidence suggests the significant role of HMGB1 in the AKI-to-CKD transition. HMGB1 is a driver of necroinflammation in AKI. Although neutralizing extracellular HMGB1 is beneficial for renal protection, HMGB1 knockdown provides additional renal protection, indicating that intracellular HMGB1 has an extracellular-independent effect[51]. Further studies have shown that intracellular HMGB1 reduces the resistance of renal tubular cells to oxidative stress[51]. The inhibition or deletion of intracellular HMGB1 promotes the proliferation and regeneration of injured renal tubular epithelial cells and reduces renal interstitial matrix deposition and neutrophil gelatinase-associated lipocalin expression, thereby improving the AKI-to-CKD transition[51]. Therefore, intracellular HMGB1 may be a potential target for enhancing kidney regeneration and improving the long-term prognosis of AKI.
4.4 Renal aging
Kidney aging increases vulnerability to disease. At the cellular level, senescence causes cells to be in a permanent and irreversible cell cycle arrest and secrete a series of proinflammatory cytokines and growth factors, known as the SASP[120]. In a D-galactose-induced age-related renal injury model, HMGB1 expression was significantly increased, accompanied by enhanced oxidative DNA damage and renal cell apoptosis[121]. HMGB1, through the NF-κB signaling pathway activation, promotes the crosstalk between the high expression of inflammatory factors and premature senescence of renal cells to play its role in renal injury[122, 123]. A recent study reported that nuclear HMGB1 directly binds to topologically associated domains or RNA to regulate proliferation or senescence [124]. In addition, HMGB1 consolidates DNA durability by increasing gaps in DNA, leading to DNA protection and improved cellular senescence[125].
4.5 Vascular calcification in CKD
Vascular calcification (VC) is an important factor contributing to CVDs-related morbidity and mortality in CKD[126]. In CKD, the accumulation of uremic toxins, oxidative stress, and chronic inflammation induces an imbalance in calcium and phosphate homeostasis and the transformation of vascular smooth muscle cells into chondrocytes or osteoblast-like cells, ultimately leading to VC[127]. HMGB1 might plausibly play a crucial role in VC in CKD. On the one hand, HMGB1 promotes osteoblastic migration and differentiation by activating RAGE/TRL4 signaling pathway[128, 129]. On the other hand, HMGB1 induces calcium deposition by regulating the expression of bone morphogenetic proteins[130]. In addition, HMGB1 reportedly initiates the mineralization process by promoting the secretion of extracellular matrix vesicles by macrophages, leading to shifted pathological mineralization[131]. Interestingly, the expression of osteopontin proteins and mineral particles promotes the cytoplasmic translocation and secretion of HMGB1[132, 133]. In a 5/6 nephrectomy-induced CKD model, a high-phosphate diet triggered inflammatory aortic calcification by promoting the nuclear-cytoplasmic translocation of HMGB1 in aortic tissue and inducing the expression of Runx2, osteopontin, and Msx2[134]. In addition, VC is also associated with the activation of Wnt/β-catenin pathway. HMGB1 promotes VC by activating the β-catenin pathway, upregulating Runx2, and downregulating Klotho in CKD[91]. Bone marrow mesenchymal stem cell-derived exosomes improve aortic calcification by promoting SIRT6 expression and reducing HMGB1 cytoplasmic translocation via deacetylation[134]. In addition, lethal giant larvae 1, a key regulator of cell polarity, can also inhibits calcification by binding to HMGB1 and promoting its degradation through the lysosomal pathway[135].
4.6 Renal replacement therapy
Renal replacement therapy or kidney transplantation is the cornerstone of patient with ESRD treatment. HMGB1 is a late inflammatory mediator in CKD. HMGB1 levels are significantly elevated in patients undergoing continuous ambulatory peritoneal dialysis (CAPD) and are associated with inflammation and malnutrition[136]. Released HMGB1 mediates peritoneal fibrosis during peritoneal dialysis (PD) treatment by promoting MCP-1 and IL-8 production[137]. Serum HMGB1 levels were significantly higher in patients undergoing hemodialysis than in patients with PD. Of note, HMGB1 levels decrease significantly with dialysis treatment[138]. Interestingly, patients with higher HMGB1 levels face more complications than those with lower HMGB1 levels despite no difference in terms of survival[138]. HMGB1 can be cleared by hemofiltration and hemodialysis using super-high-flux or high-cutoff membranes[139]. Therefore, the targeted clearance of HMGB1 by in vitro blood purification might effectively improve the clinical outcomes of critically ill patients, including ESRD[140].
5. HMGB1 antagonists of potential clinical interest in CKD
At present, several strategies have been shown to successfully inhibit HMGB1-dependent diseases, including inhibiting HMGB1 expression and release, as well as blocking HMGB1-related signaling (HMGB1/TLR4 or HMGB1/RAGE pathway) (Table 3)[7]. Targeted HMGB1 therapy has been widely studied and applied to many diseases. In CKD, Ethyl pyruvate (EP), a well-established and potent HMGB1 inhibitor, selectively inhibits HMGB1 translocation from the nucleus, which inhibits its function in the cytosol and the active secretion of HMGB1 upon cell activation[141]. EP ameliorated albuminuria and glomerular injury in an STZ-induced DKD rat model by inhibiting HMGB1[142] and alleviated CaCl2-induced renal tubular cell injury by downregulating the expression of inflammatory and autophagic proteins[143]. Glycyrrhizic acid (Gly) was the first natural HMGB1 inhibitor to be discovered. Gly induces conformational changes that interfere with the DNA-binding ability of HMGB1 in the nucleus, HMGB1 phosphorylation in the cytosol, and the binding ability of HMGB1 receptors in the extracellular space[144]. Gly ameliorates proteinuria, pathological renal injury, and disease progression in DKD rats by improving renal inflammation and ROS production by inhibiting HMGB1[145-147]. Gly can also prevent tacrolimus-induced renal injury by improving lysosomal function and regulating autophagy[148]. Many other natural products, such as Korea red ginseng[121], Bupleurum polysaccharides[149], Dioscin[150, 151], Plantago asiatica L[152], Isomangiferin[153], Troxerutin[154], and Ellagic acid[155], have been found to have similar therapeutic effects on CKD.
Therapeutic strategies targeting HMGB1 in CKD
| HMGB1 ancts | Experimental models | Mechanism | Effect on CKD | References |
|---|---|---|---|---|
| Ethyl pyruvate | STZ-induced DKD; CaCl2-induced HK-2 | Inhibit HMGB1 phosphorylation and release; Inhibit HMGB1/TLR4/NF-κB | Meliorate albuminuria and glomerular injury; prevent AKI-CKD transition | [51, 142, 143] |
| Glycyrrhizic acid | STZ-induced DKD; Zucker diabetic fatty rat | Inhibit HMGB1/RAGE/TLR4; Inhibit HMGB1/TLR4/NF-κB | Improve renal injury and inflammatory responses | [145, 146] |
| Grape seed proanthocyanidin extract | UUO | Suppress HMGB1/TLR4/p65/TGF-β1 | Alleviates renal fibrosis | [61] |
| Korea red ginseng | HFD and D-galactose-induced aging-related renal injury | Reduce extracellular HMGB1 | Restore aging-related renal injury | [121] |
| Bupleurum polysaccharides | STZ-induced DKD | Interrupt HMGB1/TLR4 | Reduce renal inflammation, fibrosis, serum creatinine level and urinary albumin excretion rate | [149] |
| Dioscin | ADR-induced renal injury; fructose-induced renal damage | Inhibit HMGB1/NF-κB | Reduce renal oxidative stress and inflammation; inhibit renal fibrosis | [150, 151], |
| Plantago asiatica L | puromycin aminonucleoside-induced renal injury | Inhibit HMGB1 | Suppress inflammation and apoptosis | [152] |
| Isomangiferin | db/db mice | Inhibit HMGB1/NLRP3/NF-κB | Inhibit renal inflammation | [153] |
| Troxerutin | methotrexate-induced nephrotoxicity | Inhibit HMGB1/RAGE/NF-κB | Inhibit inflammation and apoptosis, and activate of autophagy | [154] |
| Ellagic acid | STZ-induced DKD | Inhibit HMGB1/TLR4/NF-кB | Ameliorate oxidative renal injury | [155] |
| Anti-HMGB1 antibody | MRL/lpr lupus-prone mice; BXSB mice; cyclosporine-induced nephrotoxicity | suppress HMGB1 translocation from nuclei; Inhibit HMGB1/TLR4 | Against albuminuria; attenuate proteinuria, glomerulonephritis, circulating anti-dsDNA and immune complex deposition. | [156-158] |
| esRAGE or HMGB1 A Box | STZ-induced DKD | Block the interaction between HMGB1 and its receptors | Reduce albuminuria, glomerular injuries, interstitial fibrosis, and renal inflammation | [159] |
| Dapagliflozin | STZ-induced DKD; HG-induced HK-2; high fat diet-induced DKD | Inhibit HMGB1/TLR2/4/NF-κB; Inhibit HMGB1‑RAGE-NF‑κB | Suppress the self-perpetuating cycle of inflammation and diabetic kidney injury | [161-163] |
| Empagliflozin | STZ-induced DKD | attenuate renal HMGB1 levels | Alleviate renal inflammation and oxidative stress | [160] |
| Simvastatin | BSXSB mice | Reduce the expression of HMGB1 and TLR4 | inhibit the autoimmune response | [164] |
| Nano selenium and sildenafil | STZ-induced DKD | Inhibit HMGB1/NF-κB | Improve kidney function, and histopathological changes | [165] |
HG: high glucose; STZ: Streptozocin; HFD: High fat diet; ADR: adriamycin; UUO: unilateral ureteral obstruction.
Anti-HMGB1 antibodies have also been shown to fully inhibit the increase in complement deposition and albuminuria in MRL/lpr lupus-prone mice by inhibiting neutrophil recruitment and NETs[156, 157]. Anti-HMGB1 antibody administration inhibits NF-κB expression by blocking the activation of the TLR4 pathway, thereby improving tubulointerstitial fibrosis, improving serum creatinine and 24-hour albuminuria, reducing creatinine clearance associated with nephrotoxicity, and preventing calcineurin inhibitor-induced nephrotoxicity, which is beneficial for improving the allograft survival rate of renal transplant recipients[158]. The supra-physiological production of endogenous secretory RAGE or administration of the HMGB1 A-box also improved albuminuria, glomerular injury, interstitial fibrosis, and renal inflammation in DKD mice[159].
In addition, some drugs that have proven effective in treating CKD were found to be associated with HMGB1 inhibition. For instance, the renoprotective effect of empagliflozin alleviated renal inflammation and apoptosis and was associated with reduced levels of HMGB1, RAGE, and TLR4[160]. The renoprotective effect of dapagliflozin is related to the blocking of the renal HMGB1 feedback loop[161]. Dapagliflozin alleviates renal tubular injury, improves inflammation and oxidative stress[162], and reverses podocyte loss and fibrosis by restoring renal autophagy by inhibiting HMGB1 in DKD[163]. Simvastatin ameliorated pathological renal injury by inhibiting HMGB1 expression in the kidneys of LN mice[164].
Interestingly, with the development of computational tools, new HMGB1 inhibitors (such as nano selenium and sildenafil), have shown satisfactory effects in improving renal function and pathological damage[165]. Based on the above evidence, HMGB1 may be an attractive target for the treatment of CKD. Nevertheless, more efficient and safer HMGB1 inhibition strategies are urgently required to improve the therapeutic effects on CKD.
6. Conclusions and perspectives
HMGB1 plays multiple roles in the occurrence and progression of CKD depending on its localization, context, post-translational modification, and receptor binding. HMGB1 is expressed and secreted by stressed intrinsic renal cells and mediates renal fibrosis, aging, AKI-to-CKD transition, and cardiovascular complications by amplifying inflammation through the regulation of autophagy- and cell death-related pathways, ultimately affecting renal outcomes. In addition, as a biomarker, HMGB1 levels also significantly correlate with the progression and prognosis of CKD. Pharmacological inhibition and deletion of HMGB1 significantly improve various kidney disease phenotypes. Therefore, targeting HMGB1 is an attractive therapeutic strategy for CKD treatment. However, applying HMGB1 as a therapeutic target in CKD remains challenging. The first issue is the accurate measurement of HMGB1 levels. Studies have confirmed that HMGB1 is produced in serum during blood clots [166]. Therefore, whether plasma, serum, or urine is the best sample to predict and evaluate CKD warrants further study. In addition, studies have found that HMGB1 binds to several proteins, including IgG1, in the serum to form a complex that interferes with the enzyme linked immunosorbent assay system detection[167]. Therefore, it is important to clarify whether western blot, ELISA, liquid chromatography and tandem mass spectrometry, and other alternative methods can accurately detect HMGB1. Another critical issue that needs to be addressed is the heterogeneity in HMGB1 expression. Studies have shown sex differences in HMGB1 expression in kidney injury and that HMGB1 increases more in male rats upon kidney injury[168]. In addition, HMGB1 expression is tissue-specific[169]. Therefore, clarifying the specific factors that affect the differences in HMGB1 expression might help the development of targeted treatments for HMGB1. Finally, the decrease in renal clearance does not fully explain the increase in circulating HMGB1. Although splenectomy transiently reduced circulating HMGB1 levels and improved CKD. However, the source of HMGB1 in CKD remains to be elucidated[170]. Furthermore, at the cellular level, the dual localization of HMGB1 appears to be functionally complementary. How damaged renal intrinsic cells balance the nuclear and extracellular functions of HMGB1 remains unclear, especially, what role does intracellular HMGB1 play, and whether extracellular HMGB1 is the cause or result of kidney injury, which is the premise for identifying highly effective HMGB1 inhibitors for CKD.
Funding
This study was supported by the National Nature Science Foundation of China (82374419, 82074393, 82305210).
Competing Interests
The authors have declared that no competing interest exists.
References
1. Ruiz-Ortega M, Rayego-Mateos S, Lamas S, Ortiz A, Rodrigues-Diez RR. Targeting the progression of chronic kidney disease. Nature reviews Nephrology. 2020;16:269-88
2. KDIGO 2020 Clinical Practice Guideline for Diabetes Management in Chronic Kidney Disease. Kidney international. 2020; 98: S1-s115.
3. Lameire NH, Levin A, Kellum JA, Cheung M, Jadoul M, Winkelmayer WC. et al. Harmonizing acute and chronic kidney disease definition and classification: report of a Kidney Disease: Improving Global Outcomes (KDIGO) Consensus Conference. Kidney international. 2021;100:516-26
4. Luo S, Grams ME. Epidemiology research to foster improvement in chronic kidney disease care. Kidney international. 2020;97:477-86
5. Tang D, Kang R, Zeh HJ, Lotze MT. The multifunctional protein HMGB1: 50 years of discovery. Nature reviews Immunology. 2023
6. Chen R, Kang R, Tang D. The mechanism of HMGB1 secretion and release. Experimental & molecular medicine. 2022;54:91-102
7. Xue J, Suarez JS, Minaai M, Li S, Gaudino G, Pass HI. et al. HMGB1 as a therapeutic target in disease. Journal of cellular physiology. 2021;236:3406-19
8. Mensah MA, Niskanen H, Magalhaes AP, Basu S, Kircher M, Sczakiel HL. et al. Aberrant phase separation and nucleolar dysfunction in rare genetic diseases. Nature. 2023;614:564-71
9. Xu S, Zeng Z, Zhao M, Huang Q, Gao Y, Dai X. et al. Evidence for SIRT1 Mediated HMGB1 Release From Kidney Cells in the Early Stages of Hemorrhagic Shock. Frontiers in physiology. 2019;10:854
10. Li Y, Yuan Y, Huang ZX, Chen H, Lan R, Wang Z. et al. GSDME-mediated pyroptosis promotes inflammation and fibrosis in obstructive nephropathy. Cell death and differentiation. 2021;28:2333-50
11. Calogero S, Grassi F, Aguzzi A, Voigtländer T, Ferrier P, Ferrari S. et al. The lack of chromosomal protein Hmg1 does not disrupt cell growth but causes lethal hypoglycaemia in newborn mice. Nature genetics. 1999;22:276-80
12. Sapojnikova N, Maman J, Myers FA, Thorne AW, Vorobyev VI, Crane-Robinson C. Biochemical observation of the rapid mobility of nuclear HMGB1. Biochimica et biophysica acta. 2005;1729:57-63
13. Kwak MS, Kim HS, Lkhamsuren K, Kim YH, Han MG, Shin JM. et al. Peroxiredoxin-mediated disulfide bond formation is required for nucleocytoplasmic translocation and secretion of HMGB1 in response to inflammatory stimuli. Redox biology. 2019;24:101203
14. Yang H, Wang H, Andersson U. Targeting Inflammation Driven by HMGB1. Frontiers in immunology. 2020;11:484
15. Avalos AM, Kiefer K, Tian J, Christensen S, Shlomchik M, Coyle AJ. et al. RAGE-independent autoreactive B cell activation in response to chromatin and HMGB1/DNA immune complexes. Autoimmunity. 2010;43:103-10
16. Miao J, Ye S, Lan J, Ye P, Wen Q, Mei L. et al. Nuclear HMGB1 promotes the phagocytic ability of macrophages. Experimental cell research. 2020;393:112037
17. Dumitriu IE, Baruah P, Manfredi AA, Bianchi ME, Rovere-Querini P. HMGB1: guiding immunity from within. Trends in immunology. 2005;26:381-7
18. Deng M, Tang Y, Li W, Wang X, Zhang R, Zhang X. et al. The Endotoxin Delivery Protein HMGB1 Mediates Caspase-11-Dependent Lethality in Sepsis. Immunity. 2018;49:740-53.e7
19. Feng L, Liang L, Zhang S, Yang J, Yue Y, Zhang X. HMGB1 downregulation in retinal pigment epithelial cells protects against diabetic retinopathy through the autophagy-lysosome pathway. Autophagy. 2022;18:320-39
20. Stros M. HMGB proteins: interactions with DNA and chromatin. Biochimica et biophysica acta. 2010;1799:101-13
21. Pasheva E, Sarov M, Bidjekov K, Ugrinova I, Sarg B, Lindner H. et al. In vitro acetylation of HMGB-1 and -2 proteins by CBP: the role of the acidic tail. Biochemistry. 2004;43:2935-40
22. Bonaldi T, Talamo F, Scaffidi P, Ferrera D, Porto A, Bachi A. et al. Monocytic cells hyperacetylate chromatin protein HMGB1 to redirect it towards secretion. The EMBO journal. 2003;22:5551-60
23. Rabadi MM, Xavier S, Vasko R, Kaur K, Goligorksy MS, Ratliff BB. High-mobility group box 1 is a novel deacetylation target of Sirtuin1. Kidney international. 2015;87:95-108
24. Ito I, Fukazawa J, Yoshida M. Post-translational methylation of high mobility group box 1 (HMGB1) causes its cytoplasmic localization in neutrophils. The Journal of biological chemistry. 2007;282:16336-44
25. Youn JH, Shin JS. Nucleocytoplasmic shuttling of HMGB1 is regulated by phosphorylation that redirects it toward secretion. Journal of immunology (Baltimore, Md: 1950). 2006;177:7889-97
26. Ditsworth D, Zong WX, Thompson CB. Activation of poly(ADP)-ribose polymerase (PARP-1) induces release of the pro-inflammatory mediator HMGB1 from the nucleus. The Journal of biological chemistry. 2007;282:17845-54
27. Guha M, Saare M, Maslovskaja J, Kisand K, Liiv I, Haljasorg U. et al. DNA breaks and chromatin structural changes enhance the transcription of autoimmune regulator target genes. The Journal of biological chemistry. 2017;292:6542-54
28. Zhou J, Huang JD, Poon VK, Chen DQ, Chan CC, Ng F. et al. Functional dissection of an IFN-alpha/beta receptor 1 promoter variant that confers higher risk to chronic hepatitis B virus infection. Journal of hepatology. 2009;51:322-32
29. Davis K, Banerjee S, Friggeri A, Bell C, Abraham E, Zerfaoui M. Poly(ADP-ribosyl)ation of high mobility group box 1 (HMGB1) protein enhances inhibition of efferocytosis. Molecular medicine (Cambridge, Mass). 2012;18:359-69
30. Huang H, Nace GW, McDonald KA, Tai S, Klune JR, Rosborough BR. et al. Hepatocyte-specific high-mobility group box 1 deletion worsens the injury in liver ischemia/reperfusion: a role for intracellular high-mobility group box 1 in cellular protection. Hepatology (Baltimore, Md). 2014;59:1984-97
31. Kim J. Poly(ADP-ribose) polymerase activation induces high mobility group box 1 release from proximal tubular cells during cisplatin nephrotoxicity. Physiological research. 2016;65:333-40
32. Kim YH, Kwak MS, Park JB, Lee SA, Choi JE, Cho HS. et al. N-linked glycosylation plays a crucial role in the secretion of HMGB1. Journal of cell science. 2016;129:29-38
33. Vergoten G, Bailly C. N-glycosylation of High Mobility Group Box 1 protein (HMGB1) modulates the interaction with glycyrrhizin: A molecular modeling study. Computational biology and chemistry. 2020;88:107312
34. Balana AT, Mukherjee A, Nagpal H, Moon SP, Fierz B, Vasquez KM. et al. O-GlcNAcylation of High Mobility Group Box 1 (HMGB1) Alters Its DNA Binding and DNA Damage Processing Activities. Journal of the American Chemical Society. 2021;143:16030-40
35. Yang R, Gao Y, Li H, Huang W, Tu D, Yang M. et al. Posttranslational S-nitrosylation modification regulates HMGB1 secretion and promotes its proinflammatory and neurodegenerative effects. Cell reports. 2022;40:111330
36. Sun Y, Wang Q, Wang M, Sun F, Qiao P, Jiang A. et al. CHIP induces ubiquitination and degradation of HMGB1 to regulate glycolysis in ovarian endometriosis. Cellular and molecular life sciences: CMLS. 2022;80:13
37. Li H, Roy M, Liang L, Cao W, Hu B, Li Y. et al. Deubiquitylase USP12 induces pro-survival autophagy and bortezomib resistance in multiple myeloma by stabilizing HMGB1. Oncogene. 2022;41:1298-308
38. Zhou H, Du R, Li G, Bai Z, Ma J, Mao C. et al. Cannabinoid receptor 2 promotes the intracellular degradation of HMGB1 via the autophagy-lysosome pathway in macrophage. International immunopharmacology. 2020;78:106007
39. Volchuk A, Ye A, Chi L, Steinberg BE, Goldenberg NM. Indirect regulation of HMGB1 release by gasdermin D. Nature communications. 2020;11:4561
40. Kim YH, Kwak MS, Lee B, Shin JM, Aum S, Park IH. et al. Secretory autophagy machinery and vesicular trafficking are involved in HMGB1 secretion. Autophagy. 2021;17:2345-62
41. Yu Y, Tang D, Kang R. Oxidative stress-mediated HMGB1 biology. Frontiers in physiology. 2015;6:93
42. Wang H, Zhou XM, Wu LY, Liu GJ, Xu WD, Zhang XS. et al. Aucubin alleviates oxidative stress and inflammation via Nrf2-mediated signaling activity in experimental traumatic brain injury. Journal of neuroinflammation. 2020;17:188
43. Tsung A, Klune JR, Zhang X, Jeyabalan G, Cao Z, Peng X. et al. HMGB1 release induced by liver ischemia involves Toll-like receptor 4 dependent reactive oxygen species production and calcium-mediated signaling. The Journal of experimental medicine. 2007;204:2913-23
44. Zhao P, Ye T, Yan X, Hu X, Liu P, Wang X. HMGB1 release by H(2)O(2)-induced hepatocytes is regulated through calcium overload and 58-F interference. Cell death discovery. 2017;3:17008
45. Wu H, Xie X, Sun M, Chen M, Tao X, Fang X. et al. Modification of mesenchymal stem cells by HMGB1 promotes the activity of Cav3.2 T-type calcium channel via PKA/β-catenin/γ-cystathionase pathway. Stem cell research & therapy. 2022;13:4
46. Wu M, Gui H, Feng Z, Xu H, Li G, Li M. et al. KPT-330, a potent and selective CRM1 inhibitor, exhibits anti-inflammation effects and protection against sepsis. Biochemical and biophysical research communications. 2018;503:1773-9
47. Zhang Y, Chen Y, Zhang Y, Li PL, Li X. Contribution of cathepsin B-dependent Nlrp3 inflammasome activation to nicotine-induced endothelial barrier dysfunction. European journal of pharmacology. 2019;865:172795
48. Kuang F, Liu J, Li C, Kang R, Tang D. Cathepsin B is a mediator of organelle-specific initiation of ferroptosis. Biochemical and biophysical research communications. 2020;533:1464-9
49. Wang X, Li Z, Bai Y, Zhang R, Meng R, Chen F. et al. A small molecule binding HMGB1 inhibits caspase-11-mediated lethality in sepsis. Cell death & disease. 2021;12:402
50. Li W, Deng M, Loughran PA, Yang M, Lin M, Yang C. et al. LPS Induces Active HMGB1 Release From Hepatocytes Into Exosomes Through the Coordinated Activities of TLR4 and Caspase-11/GSDMD Signaling. Frontiers in immunology. 2020;11:229
51. Zhao ZB, Marschner JA, Iwakura T, Li C, Motrapu M, Kuang M. et al. Tubular Epithelial Cell HMGB1 Promotes AKI-CKD Transition by Sensitizing Cycling Tubular Cells to Oxidative Stress: A Rationale for Targeting HMGB1 during AKI Recovery. Journal of the American Society of Nephrology: JASN. 2023;34:394-411
52. Fan H, Tang HB, Chen Z, Wang HQ, Zhang L, Jiang Y. et al. Inhibiting HMGB1-RAGE axis prevents pro-inflammatory macrophages/microglia polarization and affords neuroprotection after spinal cord injury. Journal of neuroinflammation. 2020;17:295
53. Yang K, Fan M, Wang X, Xu J, Wang Y, Tu F. et al. Lactate promotes macrophage HMGB1 lactylation, acetylation, and exosomal release in polymicrobial sepsis. Cell death and differentiation. 2022;29:133-46
54. Personnaz J, Piccolo E, Branchereau M, Filliol A, Paccoud R, Moreau E. et al. Macrophage-derived HMGB1 is dispensable for tissue fibrogenesis. FASEB bioAdvances. 2019;1:227-45
55. Tesch G, Sourris KC, Summers SA, McCarthy D, Ward MS, Borg DJ. et al. Deletion of bone-marrow-derived receptor for AGEs (RAGE) improves renal function in an experimental mouse model of diabetes. Diabetologia. 2014;57:1977-85
56. Göcze I, Ehehalt K, Zeman F, Riquelme P, Pfister K, Graf BM. et al. Postoperative cellular stress in the kidney is associated with an early systemic γδ T-cell immune cell response. Critical care (London, England). 2018;22:168
57. Wu Y, Zwaini ZD, Brunskill NJ, Zhang X, Wang H, Chana R. et al. Properdin Deficiency Impairs Phagocytosis and Enhances Injury at Kidney Repair Phase Post Ischemia-Reperfusion. Frontiers in immunology. 2021;12:697760
58. Upadhyay R, Ying WZ, Nasrin Z, Safah H, Jaimes EA, Feng W. et al. Free light chains injure proximal tubule cells through the STAT1/HMGB1/TLR axis. JCI insight. 2020;5:e137191
59. Cheng M, Liu H, Zhang D, Liu Y, Wang C, Liu F. et al. HMGB1 Enhances the AGE-Induced Expression of CTGF and TGF-β via RAGE-Dependent Signaling in Renal Tubular Epithelial Cells. American journal of nephrology. 2015;41:257-66
60. Lynch J, Nolan S, Slattery C, Feighery R, Ryan MP, McMorrow T. High-mobility group box protein 1: a novel mediator of inflammatory-induced renal epithelial-mesenchymal transition. American journal of nephrology. 2010;32:590-602
61. Wang K, Wei H, Zhan J, Liang X, Zhang C, Liu Y. et al. GSPE alleviates renal fibrosis by inhibiting the activation of C3/ HMGB1/ TGF-β1 pathway. Chemico-biological interactions. 2020;316:108926
62. Zhang D, Wu C, Ba D, Wang N, Wang Y, Li X. et al. Ferroptosis contribute to neonicotinoid imidacloprid-evoked pyroptosis by activating the HMGB1-RAGE/TLR4-NF-κB signaling pathway. Ecotoxicology and environmental safety. 2023;253:114655
63. González-Guerrero C, Cannata-Ortiz P, Guerri C, Egido J, Ortiz A, Ramos AM. TLR4-mediated inflammation is a key pathogenic event leading to kidney damage and fibrosis in cyclosporine nephrotoxicity. Archives of toxicology. 2017;91:1925-39
64. Upadhyay R, Batuman V. Aristolochic acid I induces proximal tubule injury through ROS/HMGB1/mt DNA mediated activation of TLRs. Journal of cellular and molecular medicine. 2022;26:4277-91
65. Zmijewska AA, Zmijewski JW, Becker EJ Jr, Benavides GA, Darley-Usmar V, Mannon RB. Bioenergetic maladaptation and release of HMGB1 in calcineurin inhibitor-mediated nephrotoxicity. American journal of transplantation: official journal of the American Society of Transplantation and the American Society of Transplant Surgeons. 2021;21:2964-77
66. Rossaint J, Meersch M, Thomas K, Mersmann S, Lehmann M, Skupski J. et al. Remote ischemic preconditioning causes transient cell cycle arrest and renal protection by a NF-κB-dependent Sema5B pathway. JCI insight. 2022;7:e158523
67. Zheng S, Pan Y, Wang C, Liu Y, Shi M, Ding G. HMGB1 Turns Renal Tubular Epithelial Cells into Inflammatory Promoters by Interacting with TLR4 During Sepsis. Journal of interferon & cytokine research: the official journal of the International Society for Interferon and Cytokine Research. 2016;36:9-19
68. Garg P. A Review of Podocyte Biology. American journal of nephrology. 2018;47(Suppl 1):3-13
69. Goldwich A, Burkard M, Olke M, Daniel C, Amann K, Hugo C. et al. Podocytes are nonhematopoietic professional antigen-presenting cells. Journal of the American Society of Nephrology: JASN. 2013;24:906-16
70. Gao Z, Lu L, Chen X. Release of HMGB1 in Podocytes Exacerbates Lipopolysaccharide-Induced Acute Kidney Injury. Mediators of inflammation. 2021;2021:5220226
71. Jin J, Gong J, Zhao L, Zhang H, He Q, Jiang X. Inhibition of high mobility group box 1 (HMGB1) attenuates podocyte apoptosis and epithelial-mesenchymal transition by regulating autophagy flux. Journal of diabetes. 2019;11:826-36
72. Su Z, Li Y, Lv H, Cui X, Liu M, Wang Z. et al. CLEC14A protects against podocyte injury in mice with adriamycin nephropathy. FASEB journal: official publication of the Federation of American Societies for Experimental Biology. 2021;35:e21711
73. Abboud HE. Mesangial cell biology. Experimental cell research. 2012;318:979-85
74. Hao J, Zhang YJ, Lv X, Xu N, Liu QJ, Zhao S. et al. IFN-γ induces lipogenesis in mouse mesangial cells via the JAK2/STAT1 pathway. American journal of physiology Cell physiology. 2013;304:C760-7
75. Li R, Jia F, Ren K, Luo M, Min X, Xiao S. et al. Fibroblast growth factor inducible 14 signaling facilitates anti-dsDNA IgG penetration into mesangial cells. Journal of cellular physiology. 2021;236:249-59
76. Feng X, Yang R, Tian Y, Miao X, Guo H, Gao F. et al. HMGB1 protein promotes glomerular mesangial matrix deposition via TLR2 in lupus nephritis. Journal of cellular physiology. 2020;235:5111-9
77. Qing X, Pitashny M, Thomas DB, Barrat FJ, Hogarth MP, Putterman C. Pathogenic anti-DNA antibodies modulate gene expression in mesangial cells: involvement of HMGB1 in anti-DNA antibody-induced renal injury. Immunology letters. 2008;121:61-73
78. Alhasson F, Dattaroy D, Das S, Chandrashekaran V, Seth RK, Schnellmann RG. et al. NKT cell modulates NAFLD potentiation of metabolic oxidative stress-induced mesangial cell activation and proximal tubular toxicity. American journal of physiology Renal physiology. 2016;310:F85-f101
79. Feng X, Wu C, Yang M, Liu Q, Li H, Liu J. et al. Role of PI3K/Akt signal pathway on proliferation of mesangial cell induced by HMGB1. Tissue & cell. 2016;48:121-5
80. Wu Y, Zhao Y, Yang HZ, Wang YJ, Chen Y. HMGB1 regulates ferroptosis through Nrf2 pathway in mesangial cells in response to high glucose. Bioscience reports. 2021;41:BSR20202924
81. Dumas SJ, Meta E, Borri M, Goveia J, Rohlenova K, Conchinha NV. et al. Single-Cell RNA Sequencing Reveals Renal Endothelium Heterogeneity and Metabolic Adaptation to Water Deprivation. Journal of the American Society of Nephrology: JASN. 2020;31:118-38
82. Leung WK, Gao L, Siu PM, Lai CW. Diabetic nephropathy and endothelial dysfunction: Current and future therapies, and emerging of vascular imaging for preclinical renal-kinetic study. Life sciences. 2016;166:121-30
83. Chen J, John R, Richardson JA, Shelton JM, Zhou XJ, Wang Y. et al. Toll-like receptor 4 regulates early endothelial activation during ischemic acute kidney injury. Kidney international. 2011;79:288-99
84. Lan J, Luo H, Wu R, Wang J, Zhou B, Zhang Y. et al. Internalization of HMGB1 (High Mobility Group Box 1) Promotes Angiogenesis in Endothelial Cells. Arteriosclerosis, thrombosis, and vascular biology. 2020;40:2922-40
85. Zhou Q, Tu T, Tai S, Tang L, Yang H, Zhu Z. Endothelial specific deletion of HMGB1 increases blood pressure and retards ischemia recovery through eNOS and ROS pathway in mice. Redox biology. 2021;41:101890
86. Yu T, Xiaojuan F, Jinxi L, Xinyan M, Jie X, Yuexin T. et al. Extracellular HMGB1 Induced Glomerular Endothelial Cell Injury via TLR4/MyD88 Signaling Pathway in Lupus Nephritis. Mediators of inflammation. 2021;2021:9993971
87. Deng H, Wang C, Chang DY, Hu N, Chen M, Zhao MH. High mobility group box-1 contributes to anti-myeloperoxidase antibody-induced glomerular endothelial cell injury through a moesin-dependent route. Arthritis research & therapy. 2017;19:125
88. Zhao Z, Hu Z, Zeng R, Yao Y. HMGB1 in kidney diseases. Life sciences. 2020;259:118203
89. Chen Q, Guan X, Zuo X, Wang J, Yin W. The role of high mobility group box 1 (HMGB1) in the pathogenesis of kidney diseases. Acta pharmaceutica Sinica B. 2016;6:183-8
90. Bruchfeld A, Qureshi AR, Lindholm B, Barany P, Yang L, Stenvinkel P. et al. High Mobility Group Box Protein-1 correlates with renal function in chronic kidney disease (CKD). Molecular medicine (Cambridge, Mass). 2008;14:109-15
91. Jin X, Rong S, Yuan W, Gu L, Jia J, Wang L. et al. High Mobility Group Box 1 Promotes Aortic Calcification in Chronic Kidney Disease via the Wnt/β-Catenin Pathway. Frontiers in physiology. 2018;9:665
92. Nakamura T, Sato E, Fujiwara N, Kawagoe Y, Ueda Y, Suzuki T. et al. Positive association of serum levels of advanced glycation end products and high mobility group box-1 with asymmetric dimethylarginine in nondiabetic chronic kidney disease patients. Metabolism: clinical and experimental. 2009;58:1624-8
93. Sato F, Maruyama S, Hayashi H, Sakamoto I, Yamada S, Uchimura T. et al. High mobility group box chromosomal protein 1 in patients with renal diseases. Nephron Clinical practice. 2008;108:c194-201
94. Hossny E, El-Ghoneimy D, Soliman DA, Ashour A. Diagnostic value of serum high-mobility group box-1 in pediatric systemic lupus erythematosus. International journal of rheumatic diseases. 2019;22:1402-9
95. Abdulahad DA, Westra J, Bijzet J, Limburg PC, Kallenberg CG, Bijl M. High mobility group box 1 (HMGB1) and anti-HMGB1 antibodies and their relation to disease characteristics in systemic lupus erythematosus. Arthritis research & therapy. 2011;13:R71
96. Abdulahad DA, Westra J, Bijzet J, Dolff S, van Dijk MC, Limburg PC. et al. Urine levels of HMGB1 in Systemic Lupus Erythematosus patients with and without renal manifestations. Arthritis research & therapy. 2012;14:R184
97. Jog NR, Blanco I, Lee I, Putterman C, Caricchio R. Urinary high-mobility group box-1 associates specifically with lupus nephritis class V. Lupus. 2016;25:1551-7
98. Burbano C, Gómez-Puerta JA, Muñoz-Vahos C, Vanegas-García A, Rojas M, Vásquez G. et al. HMGB1(+) microparticles present in urine are hallmarks of nephritis in patients with systemic lupus erythematosus. European journal of immunology. 2019;49:323-35
99. Zickert A, Palmblad K, Sundelin B, Chavan S, Tracey KJ, Bruchfeld A. et al. Renal expression and serum levels of high mobility group box 1 protein in lupus nephritis. Arthritis research & therapy. 2012;14:R36
100. Wang C, Gou SJ, Chang DY, Yu F, Zhao MH, Chen M. Association of circulating level of high mobility group box 1 with disease activity in antineutrophil cytoplasmic autoantibody-associated vasculitis. Arthritis care & research. 2013;65:1828-34
101. Bruchfeld A, Wendt M, Bratt J, Qureshi AR, Chavan S, Tracey KJ. et al. High-mobility group box-1 protein (HMGB1) is increased in antineutrophilic cytoplasmatic antibody (ANCA)-associated vasculitis with renal manifestations. Molecular medicine (Cambridge, Mass). 2011;17:29-35
102. Zhu B, Li N, Zhu Q, Wu T, Heizati M, Wang G. et al. Association of serum high mobility group box 1 levels with disease activity and renal involvement in patients with systemic vasculitis. Medicine. 2019;98:e14493
103. Ma TT, Wang H, Wang C, Chang DY, Zhao MH, Chen M. Urinary levels of high mobility group box-1 are associated with disease activity in antineutrophil cytoplasmic autoantibody-associated vasculitis. PloS one. 2015;10:e0123586
104. Rayego-Mateos S, Rodrigues-Diez RR, Fernandez-Fernandez B, Mora-Fernández C, Marchant V, Donate-Correa J. et al. Targeting inflammation to treat diabetic kidney disease: the road to 2030. Kidney international. 2023;103:282-96
105. Kim J, Sohn E, Kim CS, Jo K, Kim JS. The role of high-mobility group box-1 protein in the development of diabetic nephropathy. American journal of nephrology. 2011;33:524-9
106. Nair AR, Ebenezer PJ, Saini Y, Francis J. Angiotensin II-induced hypertensive renal inflammation is mediated through HMGB1-TLR4 signaling in rat tubulo-epithelial cells. Experimental cell research. 2015;335:238-47
107. Oyama Y, Hashiguchi T, Taniguchi N, Tancharoen S, Uchimura T, Biswas KK. et al. High-mobility group box-1 protein promotes granulomatous nephritis in adenine-induced nephropathy. Laboratory investigation; a journal of technical methods and pathology. 2010;90:853-66
108. Urbonaviciute V, Voll RE. High-mobility group box 1 represents a potential marker of disease activity and novel therapeutic target in systemic lupus erythematosus. Journal of internal medicine. 2011;270:309-18
109. Zheng L, Zhang H, Tang Y. In lupus nephritis, how do extracellular DNAs trigger type I interferon secretion: Under the assistance of HMGB1-cGAS? Medical hypotheses. 2018;121:51-3
110. Wang C, Wang H, Hao J, Chang DY, Zhao MH, Chen M. Involvement of high mobility group box 1 in the activation of C5a-primed neutrophils induced by ANCA. Clinical immunology (Orlando, Fla). 2015;159:47-57
111. Li LC, Gao J, Li J. Emerging role of HMGB1 in fibrotic diseases. Journal of cellular and molecular medicine. 2014;18:2331-9
112. Yuan Y, Liu Y, Sun M, Ye H, Feng Y, Liu Z. et al. Aggravated renal fibrosis is positively associated with the activation of HMGB1-TLR2/4 signaling in STZ-induced diabetic mice. Open life sciences. 2022;17:1451-61
113. Zhou S, Yu Z, Chen Z, Ning F, Hu X, Wu T. et al. Olmesartan alleviates SARS-CoV-2 envelope protein induced renal fibrosis by regulating HMGB1 release and autophagic degradation of TGF-β1. Frontiers in pharmacology. 2023;14:1187818
114. Zheng J, Lan P, Li M, Kang MC, Xun M, Ma X. et al. Anti-Na(+)/K(+)-ATPase DR antibody attenuates UUO-induced renal fibrosis through inhibition of Na(+)/K(+)-ATPase α1-dependent HMGB1 release. International immunopharmacology. 2023;116:109826
115. Tian S, Li C, Ran R, Chen SY. Surfactant protein A deficiency exacerbates renal interstitial fibrosis following obstructive injury in mice. Biochimica et biophysica acta Molecular basis of disease. 2017;1863:509-17
116. Zhang Y. MiR-92d-3p suppresses the progression of diabetic nephropathy renal fibrosis by inhibiting the C3/HMGB1/TGF-β1 pathway. Bioscience reports. 2021;41:BSR20203131
117. Tang PM, Nikolic-Paterson DJ, Lan HY. Macrophages: versatile players in renal inflammation and fibrosis. Nature reviews Nephrology. 2019;15:144-58
118. Zhu Z, Hu J, Chen Z, Feng J, Yang X, Liang W. et al. Transition of acute kidney injury to chronic kidney disease: role of metabolic reprogramming. Metabolism: clinical and experimental. 2022;131:155194
119. Wang Z, Zhang C. From AKI to CKD: Maladaptive Repair and the Underlying Mechanisms. International journal of molecular sciences. 2022;23:10880
120. Sturmlechner I, Durik M, Sieben CJ, Baker DJ, van Deursen JM. Cellular senescence in renal ageing and disease. Nature reviews Nephrology. 2017;13:77-89
121. Park S, Kim CS, Min J, Lee SH, Jung YS. A high-fat diet increases oxidative renal injury and protein glycation in D-galactose-induced aging rats and its prevention by Korea red ginseng. Journal of nutritional science and vitaminology. 2014;60:159-66
122. Xi Y, Shao F, Bai XY, Cai G, Lv Y, Chen X. Changes in the expression of the Toll-like receptor system in the aging rat kidneys. PloS one. 2014;9:e96351
123. Chen C, Qiu R, Yang J, Zhang Q, Sun G, Gao X. et al. Lipoxin A4 Restores Septic Renal Function via Blocking Crosstalk Between Inflammation and Premature Senescence. Frontiers in immunology. 2021;12:637753
124. Sofiadis K, Josipovic N, Nikolic M, Kargapolova Y, Übelmesser N, Varamogianni-Mamatsi V. et al. HMGB1 coordinates SASP-related chromatin folding and RNA homeostasis on the path to senescence. Molecular systems biology. 2021;17:e9760
125. Yasom S, Watcharanurak P, Bhummaphan N, Thongsroy J, Puttipanyalears C, Settayanon S. et al. The roles of HMGB1-produced DNA gaps in DNA protection and aging biomarker reversal. FASEB bioAdvances. 2022;4:408-34
126. Düsing P, Zietzer A, Goody PR, Hosen MR, Kurts C, Nickenig G. et al. Vascular pathologies in chronic kidney disease: pathophysiological mechanisms and novel therapeutic approaches. Journal of molecular medicine (Berlin, Germany). 2021;99:335-48
127. Gallo Marin B, Aghagoli G, Hu SL, Massoud CM, Robinson-Bostom L. Calciphylaxis and Kidney Disease: A Review. American journal of kidney diseases: the official journal of the National Kidney Foundation. 2023;81:232-9
128. Wang B, Li F, Zhang C, Wei G, Liao P, Dong N. High-mobility group box-1 protein induces osteogenic phenotype changes in aortic valve interstitial cells. The Journal of thoracic and cardiovascular surgery. 2016;151:255-62
129. Wang B, Cai Z, Liu B, Liu Z, Zhou X, Dong N. et al. RAGE deficiency alleviates aortic valve calcification in ApoE(-/-) mice via the inhibition of endoplasmic reticulum stress. Biochimica et biophysica acta Molecular basis of disease. 2017;1863:781-92
130. Wang Y, Shan J, Yang W, Zheng H, Xue S. High mobility group box 1 (HMGB1) mediates high-glucose-induced calcification in vascular smooth muscle cells of saphenous veins. Inflammation. 2013;36:1592-604
131. Chen Q, Bei JJ, Liu C, Feng SB, Zhao WB, Zhou Z. et al. HMGB1 Induces Secretion of Matrix Vesicles by Macrophages to Enhance Ectopic Mineralization. PloS one. 2016;11:e0156686
132. Peng HH, Liu YJ, Ojcius DM, Lee CM, Chen RH, Huang PR. et al. Mineral particles stimulate innate immunity through neutrophil extracellular traps containing HMGB1. Scientific reports. 2017;7:16628
133. Passmore M, Nataatmadja M, Fung YL, Pearse B, Gabriel S, Tesar P. et al. Osteopontin alters endothelial and valvular interstitial cell behaviour in calcific aortic valve stenosis through HMGB1 regulation. European journal of cardio-thoracic surgery: official journal of the European Association for Cardio-thoracic Surgery. 2015;48:e20-9
134. Wei W, Guo X, Gu L, Jia J, Yang M, Yuan W. et al. Bone marrow mesenchymal stem cell exosomes suppress phosphate-induced aortic calcification via SIRT6-HMGB1 deacetylation. Stem cell research & therapy. 2021;12:235
135. Zhang T, Cao G, Meng X, Ouyang C, Gao J, Sun Y. et al. Lethal giant larvae 1 inhibits smooth muscle calcification via high mobility group box 1. Journal of molecular and cellular cardiology. 2020;142:39-52
136. Zhu N, Yuan W, Zhou Y, Liu J, Bao J, Hao J. et al. High mobility group box protein-1 correlates with microinflammatory state and nutritional status in continuous ambulatory peritoneal dialysis patients. Journal of artificial organs: the official journal of the Japanese Society for Artificial Organs. 2011;14:125-32
137. Chu Y, Wang Y, Zheng Z, Lin Y, He R, Liu J. et al. Proinflammatory Effect of High Glucose Concentrations on HMrSV5 Cells via the Autocrine Effect of HMGB1. Frontiers in physiology. 2017;8:762
138. Chen L, Chen G, Kong X. Serum level of high mobility group box protein-1 and prognosis of patients with end-stage renal disease on hemodialysis and peritoneal dialysis. Medicine. 2021;100:e24275
139. Naka T, Haase M, Bellomo R. 'Super high-flux' or 'high cut-off' hemofiltration and hemodialysis. Contributions to nephrology. 2010;166:181-9
140. Li Y, Chen Y, Yang T, Chang K, Deng N, Zhao W. et al. Targeting circulating high mobility group box-1 and histones by extracorporeal blood purification as an immunomodulation strategy against critical illnesses. Critical care (London, England). 2023;27:77
141. Shin JH, Kim ID, Kim SW, Lee HK, Jin Y, Park JH. et al. Ethyl pyruvate inhibits HMGB1 phosphorylation and release by chelating calcium. Molecular medicine (Cambridge, Mass). 2015;20:649-57
142. Ju KD, Shin EK, Cho EJ, Yoon HB, Kim HS, Kim H. et al. Ethyl pyruvate ameliorates albuminuria and glomerular injury in the animal model of diabetic nephropathy. American journal of physiology Renal physiology. 2012;302:F606-13
143. Zhao J, Cheng J, Li C, Xu M, Ma C, Qin L. et al. Ethyl Pyruvate Attenuates CaCl2-Induced Tubular Epithelial Cell Injury by Inhibiting Autophagy and Inflammatory Responses. Kidney & blood pressure research. 2018;43:1585-95
144. Sakamoto R, Okano M, Takena H, Ohtsuki K. Inhibitory effect of glycyrrhizin on the phosphorylation and DNA-binding abilities of high mobility group proteins 1 and 2 in vitro. Biological & pharmaceutical bulletin. 2001;24:906-11
145. Thakur V, Nargis S, Gonzalez M, Pradhan S, Terreros D, Chattopadhyay M. Role of Glycyrrhizin in the Reduction of Inflammation in Diabetic Kidney Disease. Nephron. 2017;137:137-47
146. Zhang H, Zhang R, Chen J, Shi M, Li W, Zhang X. High Mobility Group Box1 Inhibitor Glycyrrhizic Acid Attenuates Kidney Injury in Streptozotocin-Induced Diabetic Rats. Kidney & blood pressure research. 2017;42:894-904
147. Hou S, Zheng F, Li Y, Gao L, Zhang J. The protective effect of glycyrrhizic acid on renal tubular epithelial cell injury induced by high glucose. International journal of molecular sciences. 2014;15:15026-43
148. Cao R, Li Y, Hu X, Qiu Y, Li S, Xie Y. et al. Glycyrrhizic acid improves tacrolimus-induced renal injury by regulating autophagy. FASEB journal: official publication of the Federation of American Societies for Experimental Biology. 2023;37:e22749
149. Liu ZZ, Weng HB, Zhang LJ, Pan LY, Sun W, Chen HX. et al. Bupleurum polysaccharides ameliorated renal injury in diabetic mice associated with suppression of HMGB1-TLR4 signaling. Chinese journal of natural medicines. 2019;17:641-9
150. Zhang Y, Xu Y, Qi Y, Xu L, Song S, Yin L. et al. Protective effects of dioscin against doxorubicin-induced nephrotoxicity via adjusting FXR-mediated oxidative stress and inflammation. Toxicology. 2017;378:53-64
151. Qiao Y, Xu L, Tao X, Yin L, Qi Y, Xu Y. et al. Protective effects of dioscin against fructose-induced renal damage via adjusting Sirt3-mediated oxidative stress, fibrosis, lipid metabolism and inflammation. Toxicology letters. 2018;284:37-45
152. Kho MC, Park JH, Han BH, Tan R, Yoon JJ, Kim HY. et al. Plantago asiatica L. Ameliorates Puromycin Aminonucleoside-Induced Nephrotic Syndrome by Suppressing Inflammation and Apoptosis. Nutrients. 2017;9:386
153. Yue S, Xue N, Li H, Chen Z, Huang B, Wang X. Isomangiferin Attenuates Renal Injury in Diabetic Mice via Inhibiting Inflammation. Diabetes, metabolic syndrome and obesity: targets and therapy. 2020;13:4273-80
154. Arab HH, Abd El-Aal SA, Eid AH, Arafa EA, Mahmoud AM, Ashour AM. Targeting inflammation, autophagy, and apoptosis by troxerutin attenuates methotrexate-induced renal injury in rats. International immunopharmacology. 2022;103:108284
155. Zhou B, Li Q, Wang J, Chen P, Jiang S. Ellagic acid attenuates streptozocin induced diabetic nephropathy via the regulation of oxidative stress and inflammatory signaling. Food and chemical toxicology: an international journal published for the British Industrial Biological Research Association. 2019;123:16-27
156. Watanabe H, Watanabe KS, Liu K, Hiramatsu S, Zeggar S, Katsuyama E. et al. Anti-high Mobility Group Box 1 Antibody Ameliorates Albuminuria in MRL/lpr Lupus-Prone Mice. Molecular therapy Methods & clinical development. 2017;6:31-9
157. Zhang C, Li C, Jia S, Yao P, Yang Q, Zhang Y. High-mobility group box 1 inhibition alleviates lupus-like disease in BXSB mice. Scandinavian journal of immunology. 2014;79:333-7
158. Park HS, Kim EN, Kim MY, Lim JH, Kim HW, Park CW. et al. The protective effect of neutralizing high-mobility group box1 against chronic cyclosporine nephrotoxicity in mice. Transplant immunology. 2016;34:42-9
159. Chen X, Ma J, Kwan T, Stribos EGD, Messchendorp AL, Loh YW. et al. Blockade of HMGB1 Attenuates Diabetic Nephropathy in Mice. Scientific reports. 2018;8:8319
160. Ashrafi Jigheh Z, Ghorbani Haghjo A, Argani H, Roshangar L, Rashtchizadeh N, Sanajou D. et al. Empagliflozin alleviates renal inflammation and oxidative stress in streptozotocin-induced diabetic rats partly by repressing HMGB1-TLR4 receptor axis. Iranian journal of basic medical sciences. 2019;22:384-90
161. Yuan Y, Sun M, Jin Z, Zheng C, Ye H, Weng H. Dapagliflozin ameliorates diabetic renal injury through suppressing the self-perpetuating cycle of inflammation mediated by HMGB1 feedback signaling in the kidney. European journal of pharmacology. 2023;943:175560
162. Yao D, Wang S, Wang M, Lu W. Renoprotection of dapagliflozin in human renal proximal tubular cells via the inhibition of the high mobility group box 1-receptor for advanced glycation end products-nuclear factor-κB signaling pathway. Molecular medicine reports. 2018;18:3625-30
163. Feng B, Yang F, Liu J, Sun Q, Meng R, Zhu D. Amelioration of diabetic kidney injury with dapagliflozin is associated with suppressing renal HMGB1 expression and restoring autophagy in obese mice. Journal of diabetes and its complications. 2023;37:108409
164. Qin Y, Sun YT, Xia LX, Zhang YJ, Yang XJ. Effect of simvastatin on expression of IL17, HMGB1 and TLR4 in LN kidney tissues of rats. Asian Pacific journal of tropical medicine. 2014;7:792-5
165. El-Azab MF, Al-Karmalawy AA, Antar SA, Hanna PA, Tawfik KM, Hazem RM. A novel role of Nano selenium and sildenafil on streptozotocin-induced diabetic nephropathy in rats by modulation of inflammatory, oxidative, and apoptotic pathways. Life sciences. 2022;303:120691
166. Weng L, Guo L, Vachani A, Mesaros C, Blair IA. Quantification of Serum High Mobility Group Box 1 by Liquid Chromatography/High-Resolution Mass Spectrometry: Implications for Its Role in Immunity, Inflammation, and Cancer. Analytical chemistry. 2018;90:7552-60
167. Urbonaviciute V, Fürnrohr BG, Weber C, Haslbeck M, Wilhelm S, Herrmann M. et al. Factors masking HMGB1 in human serum and plasma. Journal of leukocyte biology. 2007;81:67-74
168. Mohamed R, Rafikova O, O'Connor PM, Sullivan JC. Greater high-mobility group box 1 in male compared with female spontaneously hypertensive rats worsens renal ischemia-reperfusion injury. Clinical science (London, England: 1979). 2020;134:1751-62
169. Ferrara M, Chialli G, Ferreira LM, Ruggieri E, Careccia G, Preti A. et al. Oxidation of HMGB1 Is a Dynamically Regulated Process in Physiological and Pathological Conditions. Frontiers in immunology. 2020;11:1122
170. Leelahavanichkul A, Huang Y, Hu X, Zhou H, Tsuji T, Chen R. et al. Chronic kidney disease worsens sepsis and sepsis-induced acute kidney injury by releasing High Mobility Group Box Protein-1. Kidney international. 2011;80:1198-211
171. Yang WS, Kim JS, Han NJ, Lee MJ, Park SK. Toll-like receptor 4/spleen tyrosine kinase complex in high glucose signal transduction of proximal tubular epithelial cells. Cellular physiology and biochemistry: international journal of experimental cellular physiology, biochemistry, and pharmacology. 2015;35:2309-19
172. Zhang K, Fan C, Cai D, Zhang Y, Zuo R, Zhu L. et al. Contribution of TGF-Beta-Mediated NLRP3-HMGB1 Activation to Tubulointerstitial Fibrosis in Rat With Angiotensin II-Induced Chronic Kidney Disease. Frontiers in cell and developmental biology. 2020;8:1
173. Kim SM, Lee SH, Kim YG, Kim SY, Seo JW, Choi YW. et al. Hyperuricemia-induced NLRP3 activation of macrophages contributes to the progression of diabetic nephropathy. American journal of physiology Renal physiology. 2015;308:F993-f1003
174. Chen Y, Qiao F, Zhao Y, Wang Y, Liu G. HMGB1 is activated in type 2 diabetes mellitus patients and in mesangial cells in response to high glucose. International journal of clinical and experimental pathology. 2015;8:6683-91
175. Chen B, Li Y, Liu Y, Xu Z. circLRP6 regulates high glucose-induced proliferation, oxidative stress, ECM accumulation, and inflammation in mesangial cells. Journal of cellular physiology. 2019;234:21249-59
176. Feng XJ, Liu SX, Wu C, Kang PP, Liu QJ, Hao J. et al. The PTEN/PI3K/Akt signaling pathway mediates HMGB1-induced cell proliferation by regulating the NF-κB/cyclin D1 pathway in mouse mesangial cells. American journal of physiology Cell physiology. 2014;306:C1119-28
177. Wang M, Chen X, Zhang H, Li L, Xu Y, Lu W. et al. ENSMUST00000147869 regulates proliferation and fibrosis of mesangial cells in diabetic nephropathy by interacting with Hspa9. IUBMB life. 2022;74:419-32
Author contact
![]() Corresponding authors: pengliang8028com (L. PENG); lp8675com (P. LI) and zhanyongli88com (Y. ZHAN).
Corresponding authors: pengliang8028com (L. PENG); lp8675com (P. LI) and zhanyongli88com (Y. ZHAN).