Impact Factor ISSN: 1449-2288

 Global reach, higher impact
Global reach, higher impactInt J Biol Sci 2008; 4(6):352-361. doi:10.7150/ijbs.4.352 This issue Cite
Research Paper
A PP1-binding motif present in BRCA1 plays a role in its DNA repair function
Young-Mi Yu1, Serena M. Pace1, Susan R. Allen1, Chu-Xia Deng2, Lih-Ching Hsu1,3 ![]()
1. Department of Obstetrics, Gynecology and Reproductive Sciences, School of Medicine, University of Pittsburgh, Magee-Womens Research Institute, Pittsburgh, PA15213, USA
2. Genetics of Development and Diseases Branch, National Institute of Diabetes, Digestive, and Kidney Diseases, National Institutes of Health, Bethesda, MD 20892, USA
3. School of Pharmacy, National Taiwan University College of Medicine, Taipei 10051, Taiwan, ROC
Received 2008-9-30; Accepted 2008-10-4; Published 2008-10-4
Abstract
Protein phosphatase 1α (PP1α) regulates phosphorylation of BRCA1, which contains a PP1-binding motif 898KVTF901. Mutation of this motif greatly reduces the interaction between BRCA1 and PP1α. Here we show that mutation of the PP1-binding motif abolishes the ability of BRCA1 to enhance survival of Brca1-deficient mouse mammary tumor cells after DNA damage. The Rad51 focus formation and comet assays revealed that the DNA repair function of BRCA1 was impaired when the PP1-binding motif was mutated. Analysis of subnuclear localization of GFP-tagged BRCA1 demonstrated that mutation of the PP1-binding motif affected BRCA1 redistribution in response to DNA damage. BRCA1 is required for the formation of Rad51 subnuclear foci after DNA damage. Mutation of the PP1-binding motif in BRCA1 also affected recruitment of Rad51 to sites of DNA damage. Consistent with these findings, knockdown of PP1α in BRCA1-proficient cells by small interfering RNA also significantly reduced Rad51 focus formation induced by DNA damage. Further analysis indicated that mutation of the PP1-binding motif compromised BRCA1 activities in homologous recombination. Altogether, our data implicate that interaction with PP1α is important for BRCA1 function in DNA repair.
Keywords: BRCA1, protein phosphatase 1, DNA repair
1. Introduction
Mutations of the human breast and ovarian cancer susceptibility gene BRCA1 account for the majority of familial breast and ovarian cancer cases. The BRCA1 gene encodes a nuclear phosphoprotein of 1863 amino acids (Mr ~220,000). Accumulated evidence indicates that BRCA1 plays an important role in multiple cellular functions including cell cycle checkpoint control, chromosome remodeling, transcriptional regulation, recombination, and DNA repair; therefore, is crucial for maintaining genomic integrity and chromosomal stability [1-3].
BRCA1 is expressed and phosphorylated in a cell cycle-dependent manner and is mainly phosphorylated in S phase [4,5]. BRCA1 is also phosphorylated by checkpoint kinases, including hCds1/Chk2, ATM, and ATR, in response to DNA damage induced by ionizing radiation (IR), UV or chemicals. Phosphorylation of BRCA1 occurs at multiple serine residues, such as S1524 and S1423 by ATM and ATR [6-9] and S988 by hCds1/Chk2 [10]. The phosphorylation state of BRCA1 is important for its function in G2/M checkpoint control [11] and DNA repair [12]. However, BRCA1 is primarily hypophosphorylated in mitosis and G0/G1 phases of the cell cycle [5,13], suggesting that BRCA1 must undergo dephosphorylation during G2/M transition. Whether or not dephosphorylation of BRCA1 is critical for its function remains unclear.
Serine/threonine protein phosphatase 1 (PP1) holoenzymes consist of catalytic subunits and regulatory subunits which target the catalytic subunits to distinct subcellular localizations. Together, the PP1 complexes regulate diverse cellular functions including cell cycle progression, chromosome segregation, cytokinesis, and cellular metabolism [14]. Recent studies also indicate that PP1 is involved in checkpoint activation [15] and DNA repair [16], as well as recovery from DNA damage checkpoint arrest [17]. We previously reported that the PP1α catalytic subunit interacts with BRCA1 and dephosphorylates the protein at multiple serine residues that are phosphorylated by hCds1/Chk2, ATM and ATR [18,19]. Thus, PP1α, one of the three isoforms of the PP1 catalytic subunit (α, γ, and δ), may serve as a general regulator for BRCA1 phosphorylation.
Proteins interacting with the PP1 catalytic subunit, including the majority of regulatory subunits and several substrates, share a consensus PP1-binding motif (R/K)(V/I)xF, which binds to a hydrophobic groove on the surface of the PP1 catalytic subunit and serves as an anchor for the initial binding of these proteins to PP1 [14]. Mutation of this consensus motif by substitution of phenylalanine with alanine or deletion of the motif greatly reduces the binding between these proteins and the PP1 catalytic subunit. We recently identified a PP1-binding motif 898KVTF901 in human BRCA1. eMutation of this motif substantially reduces the interaction between BRCA1 and PP1α [19].
BRCA1 is implicated in the repair of DNA double-strand breaks (DSBs) by either homologous recombination (HR) or non-homologous end-joining (NHEJ) [1,2,20]. It has been reported that BRCA1 also participates in nucleotide-excision repair [21,22]. Rad51, a homologue of bacterial RecA protein, is involved in the repair of DSBs by HR, which is an error-free DNA repair process. Expression of Rad51 is also cell cycle-dependent [23] and the protein forms nuclear foci in S phase [24]. BRCA2 has been shown to interact with Rad51 and regulate the assembly of Rad51 filaments at the sites of DNA repair. BRCA1 forms a complex with both Rad51 [25] and BRCA2 [26]. BRCA1 associates with Rad51 at subnuclear foci [25]. After DNA damage, BRCA1 is phosphorylated, relocalizes along with Rad51 to the damaged regions, and displays large nuclear foci [27,28]. BRCA1 also associates with the MRE11/RAD50/NBS1 complex, which participates in HR, NHEJ, and the DNA damage response [28]. It has been demonstrated that hCds1/Chk2 phosphorylation of BRCA1 regulates the repair of DSBs by promoting error-free HR and suppressing error-prone NHEJ [12]. BRCA1 may also participate in an error-free subtype of NHEJ [29,30]. Thus, BRCA1 is required for accurate DNA repair and BRCA1-deficient cells are hypersensitive to DNA damaging agents which induce DSBs or crosslinks of DNA strands. Here, we sought to test whether interaction with PP1 was involved in the DNA repair function of BRCA1 using Brca1∆11/∆11 mouse mammary tumor cells [31].
2. Materials and Methods
Vector construction
To construct pGFPBRCA1, a ~5.6 kb XhoI-HindIII BRCA1 cDNA fragment containing the human BRCA1 cDNA sequence (containing the stop codon) was subcloned into pGFP-C3 (Clontech, Mountain View, CA, USA). The mutant BRCA1 expression vectors, pGFPBRCA1F901A and pGFPBRCA1DEL, were constructed by replacing a 4.2 kb AflII-HindIII fragment in pGFPBRCA1 with the same fragments containing the mutations from modified pBRCA1GFPF901A and pBRCA1GFPDEL with the BRCA1 stop codon. All plasmid clones were sequence-verified.
Cell culture and transfection
A mouse mammary tumor cell line 780 (Brca1 exon 11-deleted), 293T and COS-7 cells were cultured in DMEM supplemented with 2 mM L-glutamine and 10% FBS, and ES-2 cells were cultured in McCoy's 5A medium supplemented with 1.5 mM L-glutamine and 10% FBS. Transfection was performed using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. Cells were harvested for analysis 24-48 h after transfection. Target-specific 20-25 nucleotide small interfering RNAs (siRNAs) for human PP1α and control siRNA were purchased from Santa Cruz (CA, USA) and siRNA transfection was also performed using Lipofectamine 2000.
Analysis of cell cycle distribution by flow cytometry
Cells were harvested by trypsinization, fixed in ice-cold methanol, and subjected to propidium iodide staining and flow cytometric analysis as described previously [32].
Western analysis
Cells were harvested by trypsinization and lysed in RIPA buffer supplemented with protease inhibitors. Cell lysates containing ~100 μg of protein were subjected to 4-12% SDS-PAGE, followed by Western analysis. Antibodies used were human BRCA1 antibody Ab-1 (MS110) (Calbiochem, San Diego, CA, USA), mouse Brca1 M-20, PP1α C-19, Rad51 H-92 antibodies (Santa Cruz), and γ-tubulin antibody GTU-88 (Sigma, St Louis, MO, USA). Quantitative analysis was performed using Quantity One software (BioRad, Hercules, CA, USA).
Cell survival assay
Cell survival was measured by colony formation. One day after transfection, 500 or 1000 cells were seeded in T-25 flasks. The next day, cells were either left untreated (C) or exposed to 5 Gy of IR from a 137Cs source (Sheperd Mark I 30-1 Irradiator) at a dose rate of ~110 cGy/min. Two weeks after IR treatment, colonies were fixed in cold methanol and stained with Giemsa. Colonies with at least 50 cells were then scored. Relative cell survival was calculated as the ratio of colony numbers in irradiated and untreated groups (IR/C).
Immunofluorescence staining
Cells were seeded into 4-well or 8-well chamber slides. The next day, cells were either left untreated or treated with 100 μM of 5-fluorodeoxyuridine (5-FUdR) or irradiated with 5 Gy of IR. After 24 h-treatment in 5-FUdR or 3 h, 6 h and 12 h after exposure to IR, cells were fixed on ice for 30 min in PBS-buffered 3% paraformaldehyde/2% sucrose solution and permeabilized for 5 min in cold Triton buffer (0.5% Triton X-100 in 20 mM HEPES, pH 7.4, 50 mM NaCl, 3 mM MgCl2, 300 mM sucrose)[27]. Antibody incubation was carried out as described previously [32,33]. Primary antibody used was a rabbit polyclonal anti-Rad51 (1:100 to 1:200 dilution, Santa Cruz) and secondary antibody used was FITC or Texas red-conjugated anti-rabbit at a dilution of 1:200. All slides were counter-stained with DAPI and mounted with antifade (Invitrogen). Approximately 500 cells were scored and the percentages of cells with at least ten Rad51 nuclear foci were calculated.
Comet assay
Cells were treated with IR (5 Gy) and subjected to the alkaline comet assay (single-cell gel electrophoresis) 24 h after irradiation according to the manufacturer's protocol (Trevigen, Gaithersburg , MD). Comet images were captured using a fluorescence microscope with a 20× objective (Carl Zeiss Inc., Chester, VA) and the comet tail moment (an index of DNA damage calculated as the product of tail length and fraction of DNA in the comet tail) was evaluated using Cometscore version 1.5 software (AutoComet.com). At least 50 cells were analyzed for each sample in each experiment to calculate the mean tail moment.
Homologous recombination assay
To generate cell clones carrying a substrate plasmid for the HR assay, 780 cells were transfected with 1 μg of linearized pDT220 and stable cells were selected with 5 µg/ml of puromycin. Southern blot analysis was used to identify cell clones carrying a single copy of the pDT220 substrate as described [12]. A cell clone 780pDT220, which showed two bands after EcoRV digestion (an indication of single copy insertion of pDT220), was selected for the HR assay. To determine chromosomal HR, 780pDT220 cells were co-transfected with either wild-type or mutant pBRCA1GFP and either pCMV-I-SceI or a control vector in 60-mm dishes. Cells were trypsinized and transferred to 100-mm dishes two days after transfection, and were allowed to attach to the dishes for one day. Cells were then selected with XHATM (250 µg/ml xanthine, 13.6 µg/ml hypoxanthine, 0.17 µg/ml aminopterin, 3.87 µg/ml thymidine, and 10 μg/ml mycophenolic acid)(Sigma) for three weeks. Colonies were fixed and stained with Giemsa. XHATM-resistant colonies (at least 50 cells per colony) were quantified. The HR frequency was calculated as the number of resistant colonies divided by the number of cells seeded (5 × 105 in each 100-mm dish).
Non-homologous end-joining assay
780 cells were plated in 6-well plates, and co-transfected with a linearized hyg expression vector and wild-type or mutant BRCA1GFP expression vector the next day. Cells were trypsinized and 5 × 105 cells were transferred into 100-mm dishes 24 h after transfection and subjected to selection one day later with 500 μg/ml hygromycin B for 2-3 weeks. Colonies were stained with Giemsa and counted. The NHEJ frequency was calculated as the number of resistant colonies divided by the number of cells seeded.
Statistical analysis
Data were presented as mean ± SD. Two-sided t-tests were used to assess the significance of the data. P-values < 0.05 were considered statistically significant (marked with “ * ” in the figures).
3. Results
A functional PP1-binding motif is important for BRCA1 to enhance survival of Brca1-deficient mouse mammary tumor cells after DNA damage
We previously demonstrated that transient expression of wild-type but not PP1-binding motif mutated (F901A) or deleted (DEL) human BRCA1GFP protein increased cell survival after DNA damage in a BRCA1-deficient breast cancer cell line HCC1937 [19]. Because the coding sequences for the PP1-binding motif 898KVTF901 is located in exon 11, we sought to confirm this finding in a Brca1 exon 11-deleted (Brca1∆11/∆11) mouse mammary tumor cell line 780 [31] and study the underlying mechanisms.
The full-length (FL) mouse Brca1 protein consists of 1812 amino acids [34,35]. Deletion of exon 11 leads to an in-frame loss of nucleotide sequences encoding 1105 amino acids and results in a truncated protein of 707 amino acids. As shown in Fig. 1A, 780 cells only expressed a truncated, but not a FL Brca1 protein detected by Western blot analysis using an antibody M-20, which recognizes the C-terminus of mouse Brca1. Transfection efficiency in 780 was determined using a green fluorescent protein (GFP) expression vector pEGFP-N1 and was in the range of 50-75%, which was much better than the transfection efficiency in HCC1937 cells. Expression of wild-type and mutant (F901A and DEL) BRCA1GFP proteins in 780 cells was monitored by Western analysis using a human BRCA1 antibody MS110 for every transfection experiment and the expression levels were similar (Fig. 1A). Transient expression of wild-type or mutant BRCA1GFP protein in 780 cells was consistent and did not change cell cycle distribution; whereas, protein expression in stable cell clones tended to diminish over time (data not shown). Therefore, we performed the functional assays using transiently transfected cells.
A PP1-binding motif is important for BRCA1 to enhance survival of Brca1-deficient mouse mammary tumor cells after γ-irradiation. (A) Expression of exogenous human BRCA1GFP and endogenous truncated mouse Brca1 in transiently transfected 780 cells. (B) Cell survival measured by colony formation. 780 cells were transfected with empty vector pEGFP-N1KS- (EV), wild-type pBRCA1GFP (WT), pBRCA1GFPF901A (F901A), and pBRCA1GFPDEL (DEL), followed by exposure to 5 Gy of IR. The assay was performed at least three times and similar results were obtained. * P < 0.05
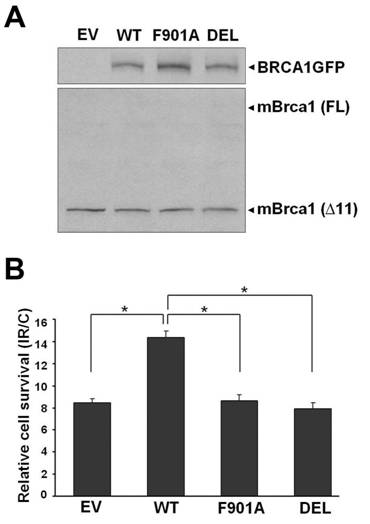
We next examined whether expression of wild-type, F901A or DEL mutant BRCA1GFP affected survival of 780 cells after exposure to γ-radiation. Transfected 780 cells were treated with 5 Gy of IR or without irradiation (C), and cell survival was measured by colony formation two weeks after γ-irradiation. As shown in Fig. 1B, relative cell survival (IR/C) was 8.5% in empty vector-transfected cells, and was significantly increased to 14.3% (a ~70% increase) in wild-type pBRCA1GFP-transfected cells (P = 0.00014, two-sided t test). Our data was comparable to the report by Scully et al., which demonstrated that expression of wild-type BRCA1 increased relative cell survival by 50-100% in HCC1937 cells exposed to 3-4 Gy of IR [36]. In contrast, expression of mutant BRCA1GFP, F901A and DEL, resulted in 8.6% and 7.9% relative cell survival respectively, which were similar to empty vector-transfected cells and significantly lower than the survival in wild-type pBRCA1GFP-transfected cells (P = 0.000295, F901A vs. wild-type; P = 0.00017, DEL vs. wild-type). Thus, consistent with our previous finding in HCC1937 cells [19], reconstitution of F901A or DEL mutant BRCA1 protein in Brca1-deficient 780 cells did not enhance survival after γ-irradiation.
Mutation of the PP1-binding motif in BRCA1 affects its DNA repair function
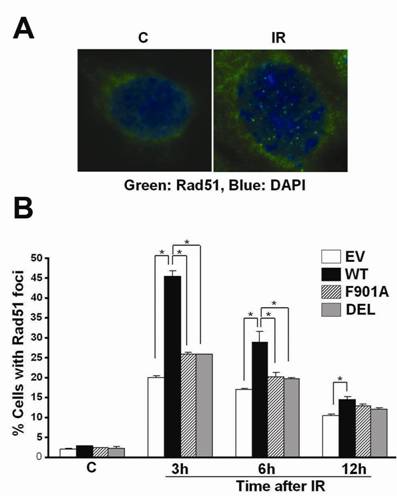
BRCA1 phosphorylation by checkpoint kinases in response to DNA damage is involved in the recruitment of DNA repair proteins including Rad51 to DNA lesions. Rad51 focus formation in the nucleus increases after DNA damage and serves as an indication of DNA repair. To determine whether the PP1-binding motif was critical for BRCA1 function in DNA repair, 780 cells were transfected with an empty vector, or pBRCA1GFP (wild-type, F901A, or DEL), left untreated (C) or treated with 5 Gy of IR, and subjected to immunofluorescence staining of Rad 51 at various time points after IR treatment. Fig. 2A shows a control cell with no Rad51 foci and an IR-treated cell with multiple nuclear Rad51 foci. Green fluorescence represents the Rad51 signal. The GFP signal of BRCA1GFP was undetectable because it was quenched in the fusion proteins. Quantitative analysis revealed that the percentage of cells with Rad51 foci (defined as cells with at least 10 Rad51 foci)[28] increased dramatically after exposure to IR, and the increase was significantly higher in wild-type pBRCA1GFP-transfected cells (45.5%) than empty vector (20.0%), pBRCA1GFPF901A (25.8%), or pBRCA1GFPDEL-transfected cells (26.0%) 3 h after γ-irradiation (Fig. 2B). The percentage of cells with Rad51 foci declined 6 h and 12 h after IR. Nevertheless, wild-type BRCA1GFP expressing cells still showed a significantly higher percentage of cells with Rad51 foci compared to others 6 h after IR. A similar trend was observed at 12 h after IR treatment, although the difference between wild-type and mutant BRCA1GFP expressing cells was not statistically significant (Fig. 2B).
Mutation of the PP1-binding motif in BRCA1 affects Rad51 focus formation after γ-irradiation. (A) A control 780 cell with no Rad51 foci and an IR-treated cell with multiple nuclear Rad51 foci. Green: Rad51, blue: DAPI counterstaining of the nucleus. (B) The percentage of transiently transfected 780 cells with Rad51 foci (defined as cells with ≥ 10 Rad51 nuclear foci) at various time points after exposure to 5 Gy of IR. EV, empty vector pEGFP-N1KS-; WT, wild-type pBRCA1GFP; F901A, pBRCA1GFPF901A; DEL, pBRCA1GFPDEL.
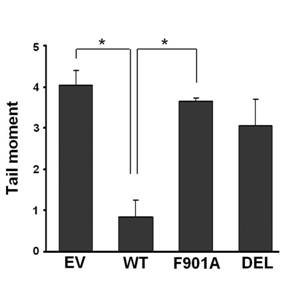
The alkaline comet assay was also performed to measure DNA strand breaks induced by IR in 780 cells expressing wild-type and mutant BRCA1GFP. DNA damage was indicated by the comet tail moment, which is the product of tail length and fraction of DNA in the comet tail. As illustrated in Fig. 3, 24 h after exposure to 5 Gy of IR, empty vector-transfected 780 cells showed a tail moment of 4.03. Expression of wild-type BRCA1GFP reduced the tail moment to 0.84 (P = 0.014). In contrast, mutant BRCA1GFP (F901A and DEL) expressing cells displayed tail moments of 3.65 and 3.06, which were not significantly different from empty vector-transfected cells. Thus, expression of wild-type but not mutant BRCA1GFP enhanced DNA repair in 780 cells.
Wild-type, but not PP1-non-binding mutant BRCA1 enhances the repair of DNA strand breaks in 780 cells determined by the comet assay. Transfected cells were exposed to 5 Gy of IR and harvested for the comet assay 24 h after irradiation.
Taken together, results from both the Rad51 focus formation assay and the comet assay provide strong evidence to support that the PP1-binding motif in BRCA1 is important for its DNA repair function.
Interaction with PP1α is important for BRCA1 to recruit Rad51 to sites of DNA damage
It has been reported that BRCA1 is required for the formation of Rad51 subnuclear foci after DNA damage [37]. We then tested whether interaction of BRCA1 with PP1α affected subcellular redistribution of BRCA1 and Rad51 after DNA damage. 5-FUdR, which showed a similar effect as IR on Rad51 focus formation, was used to induce DNA damage.
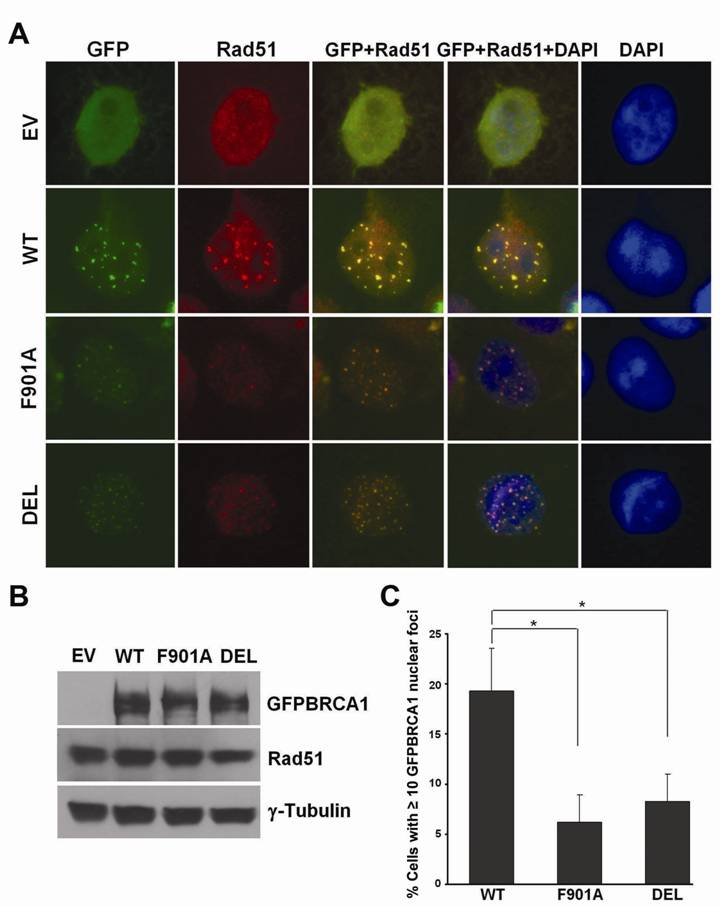
As mentioned earlier, when GFP was fused to the carboxy terminus of BRCA1, the green fluorescence of the fusion protein (BRCA1GFP) was greatly quenched. In order to improve the green fluorescent signal of the fusion proteins, we constructed a series of wild-type and mutant GFPBRCA1 expression vectors by inserting the BRCA1 coding sequences downstream from the GFP coding sequences using the pGFP-C3 vector. This series of expression vectors worked similarly as the pBRCA1GFP vectors in functional assays (data not shown). As shown in Fig. 4A, when expressed in COS-7 cells, wild-type GFPBRCA1 protein frequently formed large nuclear GFP foci after cells were treated with 100 μM of 5-FUdR for 24 h, whereas, mutant proteins usually appeared as small foci. The difference was not due to variations in GFPBRCA1 expression levels (Fig. 4B), but might be because mutant proteins were less capable to redistribute to DNA damage sites. Indeed, quantitative analysis revealed that more wild-type GFPBRCA1 expressing cells displayed 10 or more nuclear GFP foci after 5-FUdR treatment (19.1%) than mutant GFPBRCA1 expressing cells (6.7% and 8.8% respectively for F901 and DEL) as illustrated in Fig. 4C. Unlike GFPBRCA1 proteins, GFP alone did not form nuclear foci (EV in Fig. 4A). Interestingly, both wild-type and mutant GFPBRCA1 colocalized with Rad51 (Fig. 4A), suggesting that the mutation does not affect BRCA1 interaction with Rad51.
PP1-non-binding mutant BRCA1 proteins are defective in relocalization to DNA damage sites. (A) Formation of GFPBRCA1 and Rad51 nuclear foci in COS-7 cells after 5-FUdR treatment. Cells were fixed and subjected to Rad51 staining. Green: GFP or GFPBRCA1, red: Rad51. (B) Expression of GFPBRCA1 and Rad51 proteins. γ-Tubulin serves as a loading control. (C) The percentage of cells with ≥ 10 GFPBRCA1 nuclear foci induced by 5-FUdR.
We also knocked down PP1α in a BRCA1-proficient human ovarian cancer cell line ES-2 by siRNA and measured the formation of Rad51 foci after 5-FUdR treatment. Western analysis indicated that PP1α siRNA transfection resulted in more than a 50% decrease in PP1α protein level, while BRCA1 and Rad51 protein levels were not significantly affected (Fig. 5A). As shown in Fig. 5B, quantitative analysis revealed that 42.7% of control (C) siRNA-transfected cells had Rad51 foci (defined as cells with ≥ 10 Rad51 foci), whereas only 19.4% of PP1α siRNA-transfected cells displayed Rad51 foci after 5-FUdR treatment. As illustrated in Fig. 5C, PP1α siRNA transfection did not significantly change cell cycle distribution, excluding the possibility that decrease of cells with Rad51 foci by knockdown of PP1α was simply due to a cell cycle effect. Since expression of both BRCA1 and Rad51 is cell cycle-dependent and is induced in the S phase, the result from cell cycle analysis was consistent with data in Fig. 5A that showed similar expression levels of BRCA1 and Rad51 in C and PP1α siRNA-transfected cells.
Knockdown of PP1α by siRNA reduces Rad51 nuclear foci induced by 5-FUdR. (A) Expression of PP1α, BRCA1, and Rad51 in C and PP1α siRNA-transfected ES-2 cells. (B) The percentage of ES-2 cells with Rad51 foci (defined as cells with ≥ 10 Rad51 nuclear foci) induced by 5-FUdR. (C) Cell cycle distribution of C and PP1α siRNA-transfected cells.
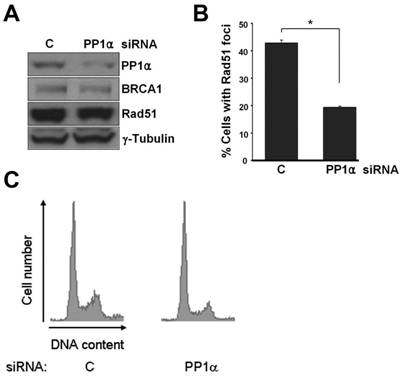
Altogether, these data indicate that interaction of BRCA1 with PP1α is important for proper relocation of BRCA1 and Rad51 to DNA damage sites, and consequently is important for the DNA repair function of BRCA1.
Promotion of homologous recombination and suppression of non-homologous end-joining by BRCA1 are partially impaired when the PP1-binding motif in BRCA1 is mutated
It has been reported that BRCA1 enhances error-free HR and inhibits error prone NHEJ to ensure accurate DNA repair [12]. Since DNA repair function of BRCA1 was impaired by mutation of the PP1-binding motif, we next assessed whether BRCA1 activities in HR and NHEJ were affected by the mutation.
To perform the HR assay, we established a stable 780 cell clone (780pDT220) with a single copy of the linearized pDT220 plasmid DNA integrated in the genome as a substrate for DSB induction and HR [12]. The recombination substrate pDT220 contains two mutated copies of the bacterial gpt gene. The upstream copy is inactivated by insertion of an I-SceI recognition site, and the downstream copy has a gene-inactivating 3' deletion in the reverse orientation. DSB induction by transient transfection of an I-SceI expression vector can lead to gene conversion that generates a functional gpt gene and confers resistance to XHATM. To assess the effect of wild-type and mutant BRCA1 on HR, 780pDT220 cells were co-transfected with pBRCA1GFP and pCMV-I-SceI. Recombinants were detected by the formation of XHATM-resistant colonies and the HR frequency was calculated as the number of resistant colonies divided by the number of cells seeded. As shown in Fig. 6A, the spontaneous HR frequency was in the range of 1.33-3.67 × 10-6 in empty vector or mutant pBRCA1GFP-transfected cells, and appeared to be higher in wild-type BRCA1GFP expressing cells at 1.57 × 10-5. Elevated spontaneous HR frequency was also observed in HCC1937 cells reconstituted with wild-type BRCA1 [12]. In the presence of DSBs generated by the I-SceI endonuclease, the HR frequency was 4.25 × 10-6 in empty vector-transfected cells, but increased dramatically to 1.05 × 10-4 in wild-type pBRCA1GFP-transfected 780 cells (P < 0.05). Expression of mutant BRCA1GFP F901A and DEL resulted in I-SceI-induced HR frequencies of 6.6 x 10-5 and 5.7 × 10-5, which were ~50% of the frequency in cells expressing the wild-type protein (P < 0.05, mutant vs. wild-type). Thus, mutation of the PP1-binding motif significantly reduced but not completely abolished the effect of BRCA1 on the promotion of I-SceI-induced HR.
The PP1-binding motif in BRCA1 compromises its DNA repair function at least in part through promoting error-free HR and to a lesser extent suppressing error-prone NHEJ. (A) HR assay. (B) NHEJ assay.
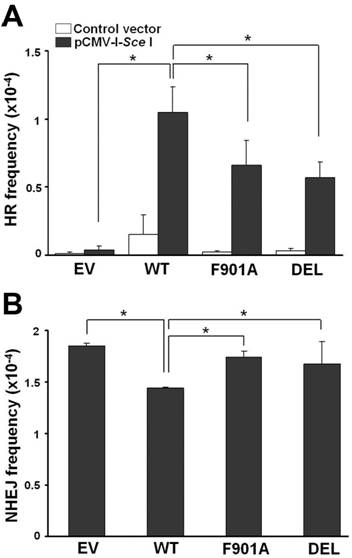
We next determined error-prone NHEJ by measuring random integration of a linearized plasmid carrying the hyg gene. 780 cells were co-transfected with the linearized plasmid and wild-type or mutant pBRCA1GFP, and subjected to hygromycin selection. Colony formation was scored 2-3 weeks after transfection. As shown in Fig. 6B, the NHEJ frequency in empty vector-transfected 780 cells was 1.85 × 10-4. The frequency was slightly suppressed with a frequency of 1.45 × 10-4 (78% of the empty vector control) when wild-type BRCA1GFP was expressed (P < 0.05). The frequencies in cells expressing mutant BRCA1GFP F901A and DEL were 1.74 × 10-4 and 1.68 × 10-4, which were statistically different from that in wild-type pBRCA1GFP-transfected cells (Fig. 6B). This result indicates that unlike in HCC1937 cells [12], BRCA1 only has a minor effect on suppressing NHEJ in 780 cells and mutation of the PP1-binding motif may also compromise this activity.
These data indicate that the PP1-binding motif in BRCA1 may play a role in enhancing DNA repair through promoting error-free HR and to a lesser extent suppressing error-prone NHEJ.
4. Discussion
Increasing evidence support the notion that BRCA1 plays a role as a sensor and/or an effector in the DNA damage repair pathway and phosphorylation of BRCA1 by checkpoint kinases is critical for this function [1,2]. Here we provide the first evidence to show that interaction with PP1α is also important for BRCA1 function in DNA repair.
In this study, we show that mutant BRCA1 proteins lacking a functional PP1-binding motif are defective in enhancing survival of Brca1-deficient mouse mammary tumor cells after DNA damage at least in part due to an impaired DNA repair function as demonstrated by the Rad51 focus formation and comet assays. We also found that PP1-binding motif-mutated BRCA1 did not relocalize as efficiently as wild-type BRCA1 to sites of DNA lesions. Consequently, the ability of BRCA1 to recruit Rad51 to the DNA lesions was also affected when the PP1-binding site was mutated. Knockdown of PP1α in BRCA1-proficient cells by siRNA also significantly reduced the percentage of cells with Rad51 foci in response to DNA damage. Altogether, these results indicate that interaction with PP1α is important for the BRCA1 function in DNA repair partly by modulating subnuclear redistribution of Rad51 after DNA damage.
BRCA1 S988 phosphorylation regulates the repair of DNA DSBs by promoting error-free HR and inhibiting error-prone NHEJ [12]. We demonstrate here that mutation of the PP1-binding motif compromises BRCA1 activities in enhancing error-free HR and to a lesser extent suppressing error-prone NHEJ assayed in Brca1-deficient mouse mammary tumor cells, which is consisitent with our previous finding that PP1α regulates S988 phsophorylation [18]. However, the effect was not as dramatic as S988A mutation in HCC1937 cells [12]. It is not clear whether the PP1-binding site in BRCA1 is also important for other DNA repair mechanisms, such as nucleotide-excision repair and error-free NHEJ. Phosphorylation of BRCA1 S1423 is important for G2/M checkpoint control [11]. Since PP1α also regulates S1423 phosphorylation [19], it is possible that mutation of the PP1-binding site may affect G2/M checkpoint function of BRCA1. This possibility is currently under investigation.
The PP1-binding motif is common to all PP1 isoforms [14]. Indeed, in vitro evidence indicates that BRCA1 also interacts with PP1γ and PP1δ (Hsu, unpublished data)[38]. Thus, it is likely that interaction with PP1, other than PP1α, is also important for BRCA1 function. Future investigation is required to provide further supporting evidence.
PP1-binding motif mutated BRCA1 is defective in DNA repair implying that dephosphorylation of BRCA1 may be required for proper DNA damage signaling or DNA repair at some point after DNA damage, emphasizing that proper phosphorylation/dephosphorylation of multiple serine residues in BRCA1 is important for BRCA1 function. Alternatively, BRCA1 may serve as a regulatory subunit for PP1 and target PP1 to other proteins whose dephosphorylation is crucial for the DNA damage repair process. These two possibilities may not be mutually exclusive, and will be investigated in our future study.
Interestingly, a missense BRCA1 mutation (K898E) has been found in the PP1-binding region. The disease relevance of this mutation is still unknown (the Breast Cancer Information Core database). Nonetheless, the presence of a germline mutation that may affect BRCA1 binding with PP1 further strengthens the biological significance of our finding. We have also found that the PPP1CA gene on human chromosome 11q13, which encodes the catalytic subunit of PP1α, is amplified and overexpressed in oral squamous cell carcinoma cell lines [39], suggesting a role of PP1α deregulation in tumor development and progression. Given that BRCA1 mutations only account for a small proportion of breast and ovarian cancers, deregulation of PP1α or other PP1 isoforms may also contribute to breast and ovarian tumorigenesis. Thus, our finding lays the foundation for additional investigation regarding the involvement of PP1 in cancer development. Furthermore, understanding the molecular mechanisms underlying the regulation of BRCA1 function by PP1 may provide new targets for the development of novel cancer therapeutics.
Acknowledgements
We thank Dr. Xia Fen for providing plasmids pDT220 and pCMV-I-SceI, Dr. Hubert Tse for help with cell irradiation, and Katherine Klingensmith for technical assistance. Young-Mi Yu was supported partly by the Magee-Womens Research Institute Postdoctoral Fellowship. This study was supported by National Cancer Institute Grant R01CA111436 and funds from the National Taiwan University (to LCH).
Conflict of Interest
The authors have declared that no conflict of interest exists.
References
1. Narod SA, Foulkes WD. BRCA1 and BRCA2: 1994 and beyond. Nat Rev Cancer. 2004;4:665-676
2. Venkitaraman AR. Tracing the network connecting BRCA and Fanconi anaemia proteins. Nat Rev Cancer. 2004;4:266-276
3. Deng CX. BRCA1: cell cycle checkpoint, genetic instability, DNA damage response and cancer evolution. Nucleic Acids Res. 2006;34:1416-1426
4. Chen Y, Farmer AA, Chen CF, Jones DC, Chen PL, Lee WH. BRCA1 is a 220-kDa nuclear phosphoprotein that is expressed and phosphorylated in a cell cycle-dependent manner. Cancer Res. 1996;56:3168-3172
5. Ruffner H, Verma IM. BRCA1 is a cell cycle-regulated nuclear phosphoprotein. Proc Natl Acad Sci USA. 1997;94:7138-7143
6. Cortez D, Wang Y, Qin J, Elledge SJ. Requirement of ATM-dependent phosphorylation of Brca1 in the DNA damage response to double-strand breaks. Science. 1999;286:1162-1166
7. Gatei M, Scott SP, Filippovitch I, Soronika N, Lavin MF, Weber B. et al. Role for ATM in DNA damage-induced phosphorylation of BRCA1. Cancer Res. 2000;60:3299-3304
8. Gatei M, Zhou B-B, Hobson K, Scott S, Young D, Khanna KK. Ataxia telangiectasia mutated (ATM) kinase and ATM and Rad3 related kinase mediate phosphorylation of Brca1 at distinct and overlapping sites. J Biol Chem. 2001;276:17276-17280
9. Tibbetts RS, Cortez D, Brumbaugh KM, Scully R, Livingston D, Elledge SJ. et al. Functional interactions between BRCA1 and the checkpoint kinase ATR during genotoxic stress. Genes Dev. 2000;14:2989-3002
10. Lee JS, Collins KM, Brown AL, Lee CH, Chung JH. hCds1-mediated phosphorylation of BRCA1 regulates the DNA damage response. Nature. 2000;404:201-204
11. Xu B, Kim S-T, Kastan MB. Involvement of Brca1 in S-phase and G(2)-phase checkpoints after ionizing irradiation. Mol Cell Biol. 2001;21:3445-3450
12. Zhang J, Willers H, Feng Z, Ghosh JC, Kim S, Weaver DT. et al. Chk2 phosphorylation of BRCA1 regulates DNA double-strand break repair. Mol Cell Biol. 2004;24:708-718
13. Thomas JE, Smith M, Tonkinson JL, Rubinfeld B, Polakis P. Induction of phosphorylation on BRCA1 during the cell cycle and after DNA damage. Cell Growth Differ. 1997;8:801-809
14. Ceulemans H, Bollen M. Functional diversity of protein phosphatase-1, a cellular economizer and reset button. Physiol Rev. 2004;84:1-39
15. Guo CY, Brautigan DL, Larner JM. Ionizing radiation activates nuclear protein phosphatase-1 by ATM-dependent dephosphorylation. J Biol Chem. 2002;277:41756-41761
16. Herman M, Ori Y, Chagnac A, Weinstein T, Korzets A, Zevin D. et al. DNA repair in mononuclear cells: role of serine/threonine phosphatases. J Lab Clin Med. 2002;140:255-262
17. den Elzen NR, O'Connell MJ. Recovery from DNA damage checkpoint arrest by PP1-mediated inhibition of Chk1. EMBO J. 2004;23:908-918
18. Liu Y, Virshup DM, White RL, Hsu L-C. Regulation of BRCA1 phosphorylation by interaction with protein phosphatase 1α. Cancer Res. 2002;62:6357-6361
19. Hsu L-C. Identification and functional characterization of a PP1-binding site in BRCA1. Biochem Biophys Res Commun. 2007;360:507-512
20. Deng C-X, Wang RH. Roles of BRCA1 in DNA damage repair: a link between development and cancer. Hum Mol Genet. 2003;12:R113-R123
21. Hartman AR, Ford JM. BRCA1 induces DNA damage recognition factors and enhances nucleotide excision repair. Nat Genet. 2002;32:180-184
22. Le Page F, Randrianarison V, Marot D, Cabannes J, Perricaudet M, Feunteun J. et al. BRCA1 and BRCA2 are necessary for the transcription-coupled repair of the oxidative 8-oxoguanine lesion in human cells. Cancer Res. 2000;60:5548-5552
23. Yamamoto A, Taki T, Yagi H, Habu T, Yoshida K, Yoshimura Y. et al. Cell cycle-dependent expression of the mouse Rad51 gene in proliferating cells. Mol Gen Genet. 1996;251:1-12
24. Tashiro S, Kotomura N, Shinohara A, Tanaka K, Ueda K, Kamada N. S phase specific formation of the human Rad51 protein nuclear foci in lymphocytes. Oncogene. 1996;12:2165-2170
25. Scully R, Chen J, Plug A, Xiao Y, Weaver D, Feunteun J. et al. Association of BRCA1 with Rad51 in mitotic and meiotic cells. Cell. 1997;88:265-275
26. Chen J, Silver DP, Walpita D, Cantor SB, Gazdar F, Tomlinson G. et al. Stable interaction between the products of the BRCA1 and BRCA2 tumor suppressor genes in mitotic and meiotic cells. Mol Cell. 1998;2:317-328
27. Scully R, Chen J, Ochs RL, Keegan K, Hoekstra M, Feunteun J. et al. Dynamic changes of BRCA1 subnuclear location and phosphorylation state are initiated by DNA damage. Cell. 1997;90:425-435
28. Zhong Q, Chen C-F, Li S, Chen Y, Wang C-C, Xiao J. et al. Association of BRCA1 with the hRad50-hMre11-p95 complex and the DNA damage response. Science. 1999;285:747-750
29. Zhang J, Powell SN. The role of the BRCA1 tumor suppressor in DNA double-strand break repair. Mol Cancer Res. 2005;3:531-539
30. Bau D-T, Mau Y-C, Shen C-Y. The role of BRCA1 in non-homologous end- joining. Cancer Lett. 2006;240:1-8
31. Brodie SG, Xu X, Qiao W, Li W-M, Cao L, Deng C-X. Multiple genetic changes are associated with mammary tumorigenesis in Brca1 conditional knockout mice. Oncogene. 2001;20:7514-7523
32. Hsu L-C, White RL. BRCA1 is associated with the centrosome during mitosis. Proc Natl Acad Sci USA. 1998;95:12983-12988
33. Hsu L-C, Doan TP, White RL. Identification of a γ-tubulin-binding domain in BRCA1. Cancer Res. 2001;61:7713-7718
34. Lane TF, Deng C, Elson A, Lyu MS, Kozak CA, Leder P. Expression of Brca1 is associated with terminal differentiation of ectodermally and mesodermally derived tissues in mice. Genes Dev. 1995;9:2712-2722
35. Marquis ST, Rajan JV, Wynshaw-Boris A, Xu J, Yin G-Y, Abel KJ. et al. The developmental pattern of Brca1 expression implies a role in differentiation of the breast and other tissues. Nat Genet. 1995;11:17-26
36. Scully R, Ganesan S, Vlasakova K, Chen J, Socolovsky M, Livingston DM. Genetic analysis of BRCA1 Function in a defined tumor cell line. Mol Cell. 1999;4:1093-1099
37. Bhattacharyya A, Ear US, Koller BH, Weichselbaum RR, Bishop DK. The breast cancer susceptibility gene BRCA1 is required for subnuclear assembly of Rad51 and survival following treatment with the DNA cross-linking agent cisplatin. J Biol Chem. 2000;275:23899-23903
38. Winter SL, Bosnoyan-Collins L, Pinnaduwage D, Andrulis IL. The interaction of PP1 with BRCA1 and analysis of their expression in breast tumors. BMC Cancer. 2007;7:85
39. Hsu L-C, Huang X, Seasholtz S, Potter DM, Gollin SM. Gene amplification and overexpression of protein phosphatase 1α in oral squamous cell carcinoma cell lines. Oncogene. 2006;25:5517-5526
Author contact
![]() Correspondence to: Dr. Lih-Ching Hsu, School of Pharmacy, National Taiwan University College of Medicine, 12F, No 1, Section 1, Jen-Ai Road, Taipei 10051, Taiwan. Phone: +886-2-2312-3456 ext. 88400; Fax: +886-2-2391-9098; E-mail: lhsuedu.tw
Correspondence to: Dr. Lih-Ching Hsu, School of Pharmacy, National Taiwan University College of Medicine, 12F, No 1, Section 1, Jen-Ai Road, Taipei 10051, Taiwan. Phone: +886-2-2312-3456 ext. 88400; Fax: +886-2-2391-9098; E-mail: lhsuedu.tw