Impact Factor ISSN: 1449-2288
- Issue 12; 2026
- Issue 11; 2026
- Issue 10; 2026
- Issue 9; 2026
- Issue 8; 2026
- Volume 22; 2026
- Past Issues
- Advance Articles
- Editorial Board
- Cover Images
- Index & Coverage
- Cover Suggestion
- Special Issues
Introduction
Hippo-YAP1/TAZ Pathway and Its...
Interaction of Hippo-YAP1/TAZ...
Interaction of Hippo-YAP1/TAZ...
Interaction of Hippo-YAP1/TAZ...
Conclusion
Acknowledgements
References

 Global reach, higher impact
Global reach, higher impactInt J Biol Sci 2020; 16(13):2454-2463. doi:10.7150/ijbs.47142 This issue Cite
Review
Role of Hippo-YAP1/TAZ pathway and its crosstalk in cardiac biology
Xiaoqing Chen1*, Wenchang Yuan2*, Yilang Li1, Jiandong Luo1 ![]() , Ning Hou1
, Ning Hou1 ![]()
1. Key Laboratory of Molecular Target & Clinical Pharmacology, School of Pharmaceutical Sciences and the Fifth Affiliated Hospital, Guangzhou Medical University, Guangzhou 511436, China;
2. KingMed School of Laboratory Medicine, Guangzhou Medical University, Guangzhou 511436, China.
*These authors contributed equally to this work.
Received 2020-4-18; Accepted 2020-6-11; Published 2020-7-6
Abstract

The Hippo pathway undertakes a pivotal role in organ size control and the process of physiology and pathology in tissue. Its downstream effectors YAP1 and TAZ receive upstream stimuli and exert transcription activity to produce biological output. Studies have demonstrated that the Hippo pathway contributes to maintenance of cardiac homeostasis and occurrence of cardiac disease. And these cardiac biological events are affected by crosstalk among Hippo-YAP1/TAZ, Wnt/β-catenin, Bone morphogenetic protein (BMP) and G-protein-coupled receptor (GPCR) signaling, which provides new insights into the Hippo pathway in heart. This review delineates the interaction among Hippo, Wnt, BMP and GPCR pathways and discusses the effects of these pathways in cardiac biology.
Keywords: Hippo pathway, Wnt signaling, BMP signaling, GPCR signaling, cardiac biology
Introduction
The Hippo pathway is a fundamental regulator that controls organ size, and first found in the Drosophila genus. Hippo, Salvador, Warts, Yorkie (Yki), and others were identified as the core components of the Hippo pathway also called Salvador-Warts-Hippo (SWH) pathway [1-3]. The Hippo pathway is highly conserved, the homologs of which are found in mammals. This pathway mediates downstream genes expression to limit organ size and take part in tumorigenesis [4]. Also, this pathway has been involved in cardiac biology including cardiogenesis, cardiac disease, and cardiac regeneration, which is mediated by cardiomyocyte proliferation, differentiation, apoptosis and others [5]. Hearts with the inactivation of Hippo pathway are subject to abnormal development, while the regeneration activity of heart and cardiomyocytes is impaired by YAP1/TAZ deficiency [6, 7]. Recent studies have revealed that multiple crosstalk among Hippo-YAP1/ TAZ, Wnt/β-catenin, Bone morphogenetic protein (BMP) and G-protein-coupled receptor (GPCR) signaling, which enriches the regulation mechanism of the Hippo pathway and the role of Hippo pathway in other signaling. In this review, we briefly recapitulate important new discoveries related to possible interactions among Hippo, Wnt, BMP and GPCR pathways, highlighting the crosstalk of these pathways in cardiac biology.
Hippo-YAP1/TAZ Pathway and Its Role in Cardiac Biology
In mammal, mammalian sterile 20-like protein kinases 1/2 (MST1/2) are the homologs of Hippo in Drosophila, phosphorylated and activated by upstream stress. Active MST1/2, together with its partner Salvador homolog 1 (SAV1), phosphorylates and activates large tumor suppressor kinase 1/2 (LATS1/2; homologs of Warts) and Mps one binder kinase activator-like 1A/1B (MOB 1A/1B; ortholog of Mats) [8, 9]. The active LATS1/2, binding to its adaptor MOB, facilitates the phosphorylation and suppression of two downstream effectors, Yes-associated protein 1 (YAP1; ortholog of Yorkie) and its paralog transcriptional coactivator with PDZ-binding motif (TAZ) (Figure 1).
The overview of Hippo-YAP1/TAZ pathway and its role in cardiac biology. The Hippo-YAP1/TAZ pathway mainly consists of MST1/2, SAV1, LATS1/2, MOB1 and two downstream effectors. The Hippo pathway, as conserved pathway in organ-size control, plays pivotal role in cardiomyocyte growth, cardiac development, cardiac disease and so on. Line arrows indicate activation, whereas dotted lines mean translocation.
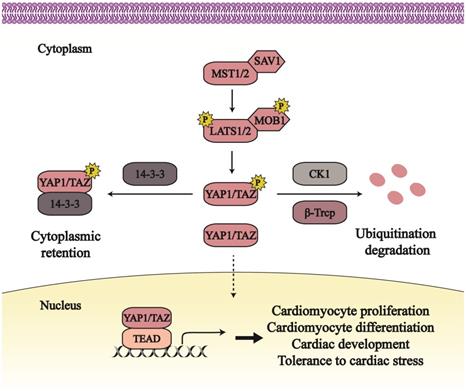
LATS1/2 can phosphorylate YAP1 at Ser61, Ser109, Ser127, Ser164, and Ser381, and target at Ser66, Ser89, Ser117, and Ser311 of TAZ [10]. While YAP1 Ser127 (Ser89 in TAZ) is phosphorylated, YAP1/TAZ binds to 14-3-3 proteins that contribute to YAP1/TAZ export from the nucleus and its retention in the cytoplasm [11]. YAP1 Ser381 phosphorylation (Ser311 in TAZ) triggers further phosphorylation by Casein kinase 1 isoform δ/ε (CK1 δ/ε), and consequently results in ubiquitination degradation dependent on the β-transducin repeat-containing E3 ubiquitin protein ligase complex (SCFβ-TRCP) [12]. YAP1/TAZ phosphorylation by the Hippo pathway has an inhibitory effect on its activity. And this inhibition can be released through Nemo-like kinase (NLK)-induced phosphorylation at YAP1 Ser128 [13].
When the Hippo pathway is in an off state, YAP1/TAZ translocates to the nucleus, where they combine with transcriptional enhanced associate domain proteins (TEADs) to facilitate gene transcription and generate downstream output. Other transcriptional partners also were found to interact with YAP1/TAZ to regulate transcription, such as Smad, and p63/p73 [14-16]. The activation of YAP1/ TAZ mainly mediates cell proliferation and apoptosis, further regulating cancer growth and organ size [4, 17].
As a highly conversed pathway, the Hippo pathway plays an essential role in controlling heart size and development [18, 19] (Figure 1). A lack of the Hippo pathway in embryonic hearts causes cardiac hypoplasia and lethality [7, 20]. SAV1 deficiency releases YAP1 from phosphorylation and then increases cardiomyocyte (CM) proliferation, which thickens ventricular walls, enlarges ventricular chambers, and leads to ventricular septal defect [7]. Mouse hearts without LATS2 or MST1/2 are characterized with the similar phenotypes. Embryos with YAP1 knockdown are hard to survive past embryonic day 16.5, subjecting to aberrant cardiac growth [21]. Except for heart growth, the Hippo signaling in adult hearts also contributes to resistance of cardiac stresses and improvement of heart survival. In response to cardiac stresses such as myocardial infraction and pressure overload, MST1 and LATS1/2 both are increased and activated to enhance CM apoptosis and reduce autophagy, subsequently leading to cardiac systolic and diastolic dysfunction [20, 22, 23]. Increasing the active form of YAP1 in the neonatal heart can rescue left ventricular tissue from myocardial infraction and improve myocardial tissue [24].
The Hippo pathway mainly mediates cell proliferation and differentiation to affect cardiac biology; YAP1/TAZ, as the downstream effectors of the Hippo pathway, facilitates the transcription of relative target genes, such as Pik3cb, Dhrs3, Sox17 and so on. Lacking of Pik3cb impedes embryonic survival; the phosphoinositide 3 kinases-protein kinase B (PI3K-AKT) signaling is activated and promotes CM proliferation via YAP1-induced Pik3cb expression [25, 26]. The differentiation activity of subepicardial cells is impaired due to enhanced Dhrs3, an inhibitor of the retinoic acid pathway [27, 28]. This may lead to the lethality of LATS1/2 cardiac knockout embryos with active YAP1 and enhanced Dhrs3 [27]. Also, YAP1 suppresses another differentiation-relative gene Sox17 that mediates the differentiation action of human embryonic stem cells [29]. CM proliferation and differentiation dependent on the Hippo pathway contribute to normal cardiac size and structure; and then YAP1/TAZ endows CMs with a primitive and fetal condition that increases CM tolerance to stresses and is beneficial to CM and heart survival.
Interaction of Hippo-YAP1/TAZ Signaling with the Wnt Pathway
The Wnt proteins can activate Wnt/β-catenin signaling (also called canonical Wnt signaling), Wnt/Ca2+ signaling, and other Wnt-related pathways. β-catenin, as an intracellular signal transducer, is pivotal in canonical Wnt signaling. In the off state, β-catenin is trapped in a destruction complex composed of AXIN, adenomatous polyposis coli (APC), Casein kinase 1 α (CK1α), and glycogen synthase kinase 3β (GSK 3β), where β-catenin is phosphorylated by CK1α and GSK 3β in turn [30]. Subsequently, the phosphorylated β-catenin suffers from ubiquitination and degradation by SCFβ-TRCP. Wnt binds to Frizzled (FZD) and Lipoprotein receptor-related protein 5/6 (LRP5/6) to initiate the Wnt signaling, consequently disassembling destruction complex [31]. β-catenin escapes from the β-catenin destruction complex into the cytoplasm and undergoes nuclear translocation to initiate target gene transcription, while other components of destruction complex are recruited to the plasma membrane. β-catenin initiates target gene transcription mostly alongside with T-cell factor/lymphoid enhancing factor (TCF/LEF) transcriptional partner [30].
Numerous studies have revealed crosstalk between the Hippo pathway and Wnt signaling (Figure 2). In canonical Wnt/β-catenin signaling, CK1ε (another member of the CK1 family) combines with and activates dishevelled 2 (DVL2) to transduce biological signals from the Wnt receptors. They, as positive regulators, destabilize β-catenin degradation complex and recruit the complex to cell membrane in order to reduce β-catenin degradation and promote its activity [32, 33]. MST1/2, a core serine/threonine kinase in the Hippo pathway, inhibits canonical Wnt/β-catenin signaling independent on YAP1/TAZ [34]. MST1/2 binds to CK1ε, subsequently disrupting the relationship between DVL2 and CK1ε as well as impeding the CK1ε-induced phosphorylation and activation of DVL2 [34]. This MST1/2-CK1ε interaction suppresses cell proliferation, which can be recovered by Wnt3a treatment. Expect for MST1/2, YAP1/TAZ also can disturb CK1ε-DVL2 binding. Phosphorylated TAZ combines with DVL2 and therefore inhibits the interaction of DVL2 and CK1ε; TAZ deficiency in mouse enhances nuclear β-catenin and target gene transcription [35]. SAV1, another member of Hippo pathway, was found to inactive β-catenin through the Hippo pathway. β-catenin activation in the heart of SAV1-deficient embryos enhances cell proliferation, expansion of ventricular myocardial layers, and thickening of ventricular walls, whereas β-catenin deficiency reduces SAV1 CKO-induced cardiac expansion [7]. Therefore, active Hippo-YAP1/TAZ pathway dampens Wnt/β-catenin signaling and its proliferation effects in cardiac growth.
The crosstalk between the Hippo-YAP1/TAZ pathway and Wnt signaling. The Wnt/ β-catenin signaling and alternative Wnt signaling participate in the modulation of the Hippo-YAP1/TAZ pathway. Conversely, the Hippo-YAP1/TAZ pathway plays a role in the regulation of Wnt signaling. The crosstalk between the Hippo pathway and Wnt signaling affects cancer biological behavior, cardiomyocyte proliferation and cardiac growth. Line arrows indicate activation, whereas connector lines imply inhibition; dotted lines mean translocation.
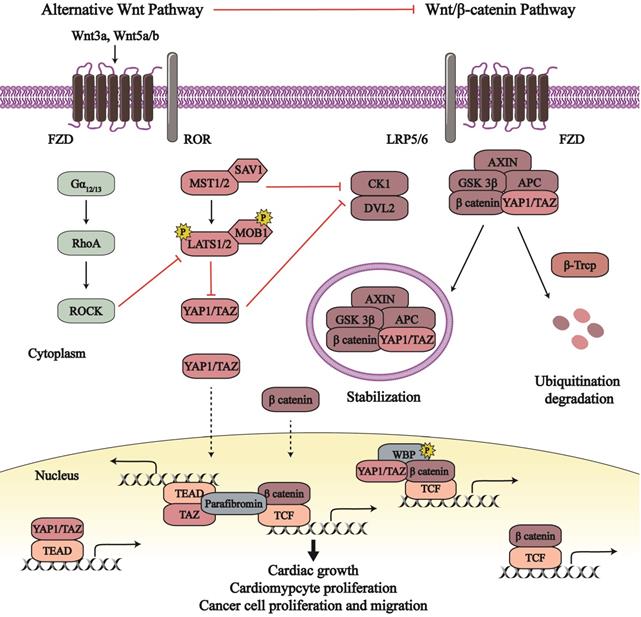
In basal epidermal cells, YAP1-5SA, an active form of YAP1, overexpression enhances active β-catenin in the nucleus, leading to enhanced cell proliferation [36]. As mentioned in section 2, YAP1 can upregulate Pik3bc and then activate PI3K-AKT pathway in CM [25]. YAP1-S112A (constitutively active YAP1) inhibits GSK 3β activity in the embryonic heart by activating the insulin-like growth factor-PI3K/AKT pathway; subsequently, reduced phosphorylation of β-catenin by GSK 3β leads to enhanced β-catenin activity [25, 37]. β-catenin undertakes positive mediators in CM proliferation [38]. As result, YAP1 may indirectly mediate Wnt/β-catenin signaling via enhancing the activity of PI3K/AKT pathway, which can affect CM proliferation. Moreover, YAP1-induced activation of canonical Wnt/β-catenin signaling may rescue AC16 human CMs after ischemia-reperfusion (IR) injury [39]. So, YAP1-dependent β-catenin activation results in the promotion of CM proliferation and survival, contributing to cardiac growth. And YAP1 and β-catenin synergistically increase CM proliferation by respective transcriptional target genes, which mediate cardiac development and heart diseases.
Recent reports have found that YAP1/TAZ is combined into the β-catenin degradation complex, which affects Wnt/β-catenin pathway and the Hippo pathway at the same time. In the absence of Wnt, YAP1/TAZ binds to the β-catenin degradation complex and the latter recruit SCFβ-TRCP for the degradation and devitalization of YAP1/TAZ and β-catenin [40, 41]. Following Wnt ligand stimulation, both YAP1/TAZ and β-catenin escape from the disassemble degradation complex, subsequently translocate to the nucleus, and interact with different transcription factors (such as TEADs and TCF/LEF) to produce respective downstream effects. What's more, the YAP1/TAZ-β-catenin destruction complex can be stranded in the multi-vesicular body compartment to protect themselves against degradation, as so to exert their biological output [42]. Except for combining into the β-catenin degradation complex, cytoplasmic YAP1/TAZ directly binds to β-catenin and retains the latter in the cytoplasm to inhibit β-catenin nuclear accumulation [43]. This direct binding is conducive to balance between Hippo pathway and Wnt/β-catenin signaling in neurogenesis and tumorigenesis [43, 44]. In the nucleus, YAP1/TAZ and β-catenin bind to not only their respective transcription factors but also each other to stimulate transcriptional actions. With the Wnt pathway in the on state, the YAP1/TAZ-β-catenin complex combines with the phosphorylated WW domain-binding protein 2 (WBP2), enters the nucleus, and provokes transcriptional action by the β-catenin-TCF complex [45]. Besides, the TAZ-β-catenin complex associates with parafibromin (a nuclear scaffold protein) which cooperatively initiates TEAD and TCF co-activator activities [46].
Interestingly, a growing body of evidence indicates that Wnt proteins can activate YAP1/TAZ, independent of Wnt/β-catenin signaling in many cell lines [47-51]. An alternative Wnt pathway has been promulgated, consisting of Wnt, FZD/ROR, Ga12/13, Rho GTPases, LATS1/2, and YAP1/TAZ, wherein YAP1 and TAZ are the key downstream transcription cofactors [49]. The Wnt5a/b and/or Wnt3a, as extracellular stimulants, activate FZD-receptor tyrosine kinase-like orphan receptor 1/2 (ROR1/2) receptor complex to transmit biological signals. Therewith, this stimulates the inhibitory effect of the Ga12/13-Rho GTPases axis in LATS1/2, which increases the nuclear sequestration and activity of YAP1/TAZ. Furthermore, Wnt5a/b are identified as target genes of YAP1/TAZ, which suggests the existence of a positive feedback loop in alternative Wnt-YAP1/TAZ signaling to reinforce alternative Wnt pathway [49].
This fascinating axis regulates several biological effects, including osteogenic differentiation, cell migration, cell polarity, and cytoskeletal oscillation. Wnt3a activates the Wnt-YAP1/TAZ pathway to maintain the pluripotency of bovine trophoblast stem cells (TSCs) [50]. The Wnt5a-induced YAP1/TAZ activation mediates cell polarity and formation of organ primordia [47]. Wnt-activated YAP1 regulates rostral and caudal brain tissue identity and affects neural tissue from human-induced pluripotent stem cells (hiPSCs) [51]. However, there are no more studies about the relationship between Wnt-YAP1/TAZ pathway and cardiac biology.
When the alternative Wnt-YAP1/TAZ pathway is activated, there is increased expression of Wnt/β-catenin signaling inhibitors including Wnt5a, Dickkopf-related protein 1 (DKK1), BMP4, connective tissue growth factor (CTGF), and cysteine-rich angiogenic inducer 61 (CYR61), all of which attenuate canonical Wnt/β-catenin signaling [49, 52]. However, the afore-mentioned studies describe that active YAP1/TAZ can boost β-catenin activity in canonical Wnt signaling, and directly participate in β-catenin transcriptional events as a co-transcriptional factor. This controversy implies more detailed crosstalk between the Hippo-YAP1/TAZ pathway and Wnt/β-catenin signaling that needs further investigation to understand this sophisticated regulatory system.
Interaction of Hippo-YAP1/TAZ Signaling with the BMP Pathway
Bone morphogenetic proteins (BMPs) belong to the family of transforming growth factor-β (TGF-β) and are involved in embryonic development as well as organ and tissue homeostasis. BMPs associate with type 1 and 2 cell membrane receptors to activate and phosphorylate intercellular receptor-regulated Smads (Smad1, Smad5, and Smad8) that combine with common partner Smads (Smad4) and accumulate in the nucleus [53]. The Smads complex binds with transcriptional factors to regulate target gene expression [54]. The BMP pathway and Hippo signaling pathway intersect to regulate embryonic growth and maintain cell homeostasis (Figure 3).
The crosstalk between the Hippo-YAP1/TAZ pathway and BMP signaling. The Hippo-YAP1/TAZ pathway bidirectionally regulate the bone morphogenetic proteins (BMP) pathway. YAP1/TAZ binding to Smads form transcription complex to regulate respective output in cancer cells, embryonic stem cells (ESC), endothelial cells as well as heart. Line arrows indicate activation, whereas connector lines imply inhibition; dotted lines mean translocation.
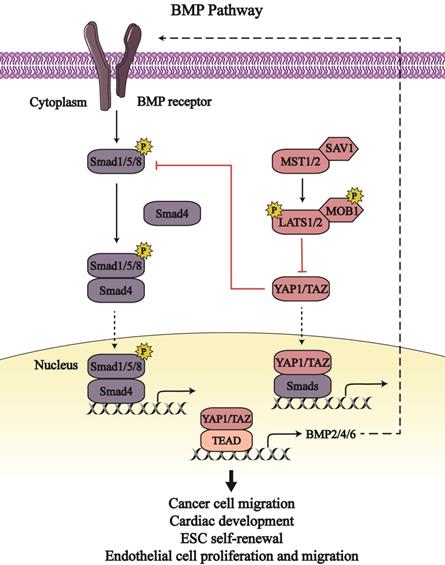
In Drosophila, the transcription activity of the Smads complex is potentiated due to impaired Yki activity, which elicits defective escort-cell germline differentiation [55]. In human and mouse mesenchymal stromal/stem cells (MSCs), active YAP1 inhibits the phosphorylation of Smad1, Smad5, and Smad8, and downregulates the expression of BMP signaling target genes, such as inhibitor of DNA-binding proteins (Id1, Id2, and Id3) [56]. The BMP-induced Ids output can regulate cell differentiation; definitively, Id1 can restrain the differentiation and sustain self-renewal activity of embryonic stem cells (ESC) [57]. So, YAP1 may downregulate Ids expression via BMP signaling as so to affect embryonic self-renewal capacity. Moreover, YAP1/TAZ inhibits BMP signaling through the augmented expression of BMP antagonists, such as CTGF, cysteine-rich motor neuron 1 protein (CRIM1), Follistatin, BMP and activin membrane-bound inhibitor homolog (BAMBI), E3 ubiquitin-protein ligase SMURF1/2, and Noggin [58, 59]. Thus, YAP1/TAZ enhances the proliferation, migration and barrier function of endothelial cells to prevent bleeding in response to mechanical stretch by upregulating BMP antagonists as well as downregulating BMP signaling [58].
Several BMPs have been identified as target genes of YAP1/TAZ, including BMP2, BMP4 and BMP6. YAP1/TAZ can enhance BMP2 transcription and expression that are inhibited by MOB1/2 and LATS1/2 [60, 61]. Enhanced BMP2 impairs the function and homeostasis of intestinal epithelial cells [61]. Moreover, BMP2 induces epithelial-to-mesenchymal transition (EMT) in the cardiac cushion, which is conducive to the development of valve and septa [54]. YAP1-mediated BMP2 expression results in the generation of the venous pole contributing to the growth of heart atrium [60]. BMP4 is upregulated by YAP1 transcription coactivator in zebrafish and mouse endothelial cells [62, 63]. Following BMP4 stimulation, active BMP signaling induces the apoptosis of human umbilical vein endothelial cells and dampens endothelial tubulogenesis [64]. However, BMP4 treatment enhances the proliferation and migration of mouse embryonic endothelial cells and human microvascular endothelial cells via activation of vascular endothelial growth factor/vascular endothelial growth factor receptor 2 (VEGF/VEGFR2) signaling [65]. These differences of BMP signaling in endothelial cells may be caused by the different downstream signaling of BMP4 and cell lines used. Also, YAP1-dependent BMP6 expression contributes to endothelial cell migration and tube formation via activating BMP pathway [66]. As such, the Hippo pathway stimulates BMP signaling by increasing the content of BMPs, which maintains endothelial cell functions and cardiac development.
Hereditary hemorrhagic telangiectasia (HHT) is a vascular syndrome characterized by vascular malformations [67]. Among the five subtypes of HHT, HHT5 is caused by GDF2 (encoding BMP9) mutations [68]. In response to BMP9 stimulation, YAP1 translocates to the nucleus to form a complex with Smad1/5, and facilitates CTGF/CYR61 transcription in endothelial cells; this mechanism partly explains the etiology of HHT [69]. Except for Smad1/5, YAP1/TAZ can bind to Smad2/3 in the nucleus to enhance Smad2/3 nuclear accumulation, which improves BMP signaling activity and EMTs [69, 70]. Notably, YAP1 and BMP are both activated in the neural crest of avian embryos; active YAP1 is indispensable for BMP activity, whereas BMP is sufficient for YAP1 activity in neural crest migration [71]. To the extent, there exists a positive and bidirectional interaction between BMP pathway and YAP1/TAZ.
Interaction of Hippo-YAP1/TAZ Signaling with the GPCR Pathway
G-protein-coupled receptors (GPCRs) are a type of cell-surface transmembrane receptors that contact intercellular G proteins to regulate second messengers to elicit downstream effects [72], wherein the Hippo-YAP1/TAZ signaling pathway has garnered attention (Figure 4).
The crosstalk between the Hippo-YAP1/TAZ pathway and GPCR signaling. G-protein-coupled receptors (GPCR) signaling regulate the Hippo-YAP1/TAZ pathway by serval G proteins and different downstream signaling. Gα12/13, Gαq/11, and Gαi, together with RhoA-ROCK axis and PKs axis, mainly upregulate YAP1/TAZ; while Gαs generally inhibits YAP1/TAZ by PKA axis. GPCR signaling-dependent Hippo pathway regulation affects the proliferation, migration and survival of cancer cells, endothelial cells and cardiomyocytes. Line arrows indicate activation, whereas connector lines imply inhibition; dotted lines mean translocation.
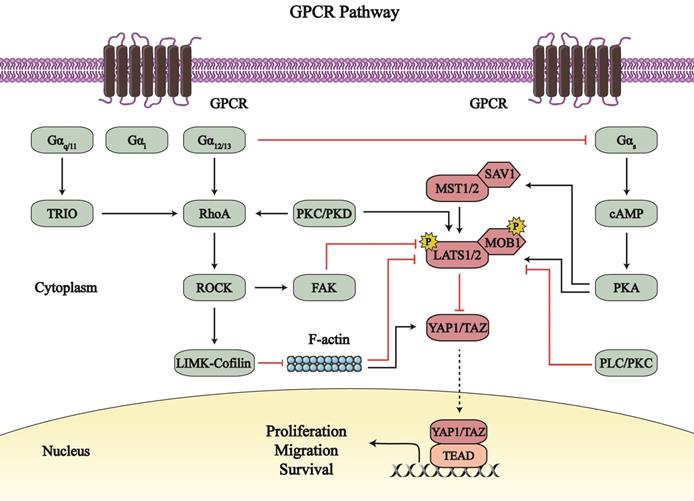
The Hippo-YAP1/TAZ pathway is regulated by different heterotrimeric G proteins. YAP1/TAZ is generally activated by Gα12/13, Gαq/11, and Gαi; these G proteins are coupled with different GPCRs, including lysophosphatidic acid receptors, sphingosine1-phosphate (S1P) receptors, and protease-activated receptor 1 [73-79]. However, Gαs inhibits the Hippo-YAP1/TAZ pathway by other GPCRs, such as free fatty acid receptor 1/4 (FFAR1/4, also called as G-protein receptor 40/120 [GPR40/120]) [80-82].
Gα12/13, Gαq/11, and Gαi mainly upregulate Ras homology family member A (RhoA)-Rho associated protein kinase (ROCK) signaling to catalyze the phosphorylation of its substrates, which affects YAP1/TAZ. The LATS1/2-induced phosphorylation of YAP1/TAZ is suppressed by the Gα12/13-RhoA-ROCK pathway, with the net effect of increased YAP1/TAZ nuclear location and activity [73, 74]. Activated YAP1 potentiates the transcription and expression of the glycolytic enzyme phosphoglycerate mutase (PGAM1) as well as c-Myc in osteosarcoma cells, which may contribute to the reprogramming of glucose metabolism in the tumor [74]. In addition to canonical Hippo signaling, the Gα12/13-RhoA-ROCK axis regulates F-actin and myosin assembly to activate YAP1/TAZ [78, 79, 83]. Specifically, the G-protein-coupled estrogen receptor 1 (GPER; also known as GPR30) acts via the Rho-ROCK signaling to mediate the LIM-domain kinase (LIMK)-Cofilin pathway that stabilizes F-actin and promotes cytoskeleton assembly; both F-actin and myosin activation activates YAP1/TAZ, independent on LATS1/2 inhibition [83]. YAP1/TAZ activation by the Gα12/13-RhoA-ROCK axis can promote endothelial cell sprouting, facilitate vascular smooth muscle cell (VSMC) migration and proliferation, and may participate in embryonic vascular development [73]. Furthermore, Rho-ROCK axis promote trophectoderm differentiation and inhibit the formation of the inner cell mass through enhanced YAP1 nuclear localization [84]. Thus, the Gα12/13-RhoA-ROCK axis may participate in YAP1/TAZ-mediated cardiac biology.
Likewise, Gαq/11, and Gαi mediate RhoA-ROCK signaling to affect the Hippo pathway. The Gαq activates TRIO-RhoA asix (non-canonical Gαq-signaling) in uveal melanoma. The Gαq-TRIO-RhoA axis promotes focal adhesion kinase (FAK)-regulated MOB1 Tyr26 phosphorylation, which disrupts the inhibition of the MOB1-LATS complex on YAP1; simultaneously, this axis enhances YAP1 Tyr357 phosphorylation to activate YAP1 and facilitate cell growth [75]. The S1P-S1P receptor (S1PR) pathway acts via Gαi to stabilize YAP1 and promote the survival of pancreatic progenitor cells [85]. However, the results about S1PR-induced YAP1 activation are different from reports from Kemppainen Lab. The S1PR2-Gα12/13 axis stimulated YAP1 and enhanced its transcriptional activity in response to 1 h treatment with sphingosylphosphorylcholine (SPC); however, longer SPC treatment (9 h) led to higher levels of phosphorylated YAP1 Ser127, and lower levels of its target genes [86]. With short SPC treatment, S1PR2 may activate Gαq, Gα12/13, and Gαi subunits to dephosphorylate and stabilize YAP1 [85, 86]. During long SPC treatment, S1PR2 may indirectly affect the Gαs downstream pathway via Gα12/13, so that LATS1/2 is activated and phosphorylated YAP1 Ser127 is increased [86, 87]. And there exists a negative feedback mechanism that YAP1/TAZ increase endogenous LATS2 activity through direct induction of TEAD-dependent Lats2 transcription [88]. These special mechanisms may lead to the long SPC treatment-induced suppression of YAP1.
Except for RhoA- ROCK axis, other signaling also mediates YAP1/TAZ. In triple-negative breast cancer (TNBC) cells, bisphenol S impacts the GPER-activated phospholipase C/protein kinase C (PLC/PKC) pathway, which decreases the levels of phosphorylated-LATS1/2 and subsequently increases YAP1 dephosphorylation [89]. Therefore, PLC/PKC-mediated YAP1 activation promotes TNBC cells migration. P2Y2 receptor (P2Y2R) is a purinergic GPCR that regulates Gαq/11-PKC signaling to control the flow of Ca2+ and K+ ions [90]. The P2Y2R-PKC axis activates YAP1 by inhibiting LATS1/2 and subsequently promote cell proliferation in human c-Kit+ cardiac progenitor cells (hCPCs), which helps to rescue CM from damages [91, 92]. Moreover, the GPCR agonist neurotensin attenuates YAP1 phosphorylation at Ser127/397 and augments YAP1 nuclear localization via PKD signaling [93]. Most importantly, PK-induced YAP1/TAZ activation is divided into two phases [94]. In the first phase, the GPCR-PKC/PKD axis induces rapid activation of LATS1/2, to enhance the phosphorylation of YAP1 at Ser127. Next, the GPCR-PKC/PKD axis activates RhoA and induces the formation of the actin cytoskeleton, and subsequently dephosphorylates YAP1 and promotes YAP1-TEAD activity.
Paradoxically, PKs, together with Gαs, impairs the activity of YAP1/TAZ. In combination with Gαs, FFAR1/2/4 activates PKA-induced phosphorylation of MST1/LATS1; thereafter, YAP1 is phosphorylated and detained in the cytoplasm, which inhibits cell proliferation and cell metastasis, and induces apoptosis of tumor cells [80-82]. Notably, the Gαs-PKA-induced suppression of YAP1 can be disrupted by positive G proteins, including Gα12/13, Gαq/11, and Gαi [95]. GPCRs exert bidirectional influence on the Hippo-YAP1/TAZ pathway, which is mutually restricted. However, the relationship between GPCRs and the Hippo-YAP1/TAZ pathway in cardiac biology is unclear.
Conclusion
Accumulating reports focus on the Hippo-YAP1/TAZ signaling in cardiac biology. As an important heart-control signaling, the Hippo pathway dysregulation contributes to abnormal cardiac development and cardiac diseases. In this review, we discuss the crosstalk between Hippo pathway and other signaling including Wnt/β-catenin, BMP and GPCR signaling. Wnt signaling and GPCR signaling mainly act as the upstream regulation of the Hippo-YAP1/TAZ pathway, affecting the activity of its core members as well as directly mediating YAP1/TAZ subcellular location and abundance. The Hippo signaling pathway also controls the output of BMP signaling and Wnt/β-catenin signaling by regulating their kinases activity and transcriptional events. Such, the interaction among these pathways may result in the role of the Hippo pathway in cardiac biology, which may explain the complex output of Hippo pathway in heart and cardiomyocytes. Accordingly, the crosstalk between Hippo pathway and other signaling sheds light on physiological events of cardiac development and pathophysiological mechanism of cardiac diseases.
Acknowledgements
This work was supported by the National Natural Science Foundation of China [No. 81402928, 81773720] to Dr. Ning Hou.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Halder G, Johnson RL. Hippo signaling: growth control and beyond. Development. 2011;138:9-22
2. Xu T, Wang W, Zhang S, Stewart RA, Yu W. Identifying tumor suppressors in genetic mosaics: the Drosophila lats gene encodes a putative protein kinase. Development. 1995;121:1053-63
3. Pan D. The hippo signaling pathway in development and cancer. Dev Cell. 2010;19:491-505
4. Mo J-S. The role of extracellular biophysical cues in modulating the Hippo-YAP pathway. BMB Reports. 2017;50:71-8
5. Zhou Q, Li L, Zhao B, Guan KL. The hippo pathway in heart development, regeneration, and diseases. Circ Res. 2015;116:1431-47
6. Heallen T, Morikawa Y, Leach J, Tao G, Willerson JT, Johnson RL. et al. Hippo signaling impedes adult heart regeneration. Development. 2013;140:4683-90
7. Heallen T, Zhang M, Wang J, Bonilla-Claudio M, Klysik E, Johnson RL. et al. Hippo pathway inhibits Wnt signaling to restrain cardiomyocyte proliferation and heart size. Science. 2011;332:458-61
8. Chan EH, Nousiainen M, Chalamalasetty RB, Schafer A, Nigg EA, Sillje HH. The Ste20-like kinase Mst2 activates the human large tumor suppressor kinase Lats1. Oncogene. 2005;24:2076-86
9. Praskova M, Xia F, Avruch J. MOBKL1A/MOBKL1B phosphorylation by MST1 and MST2 inhibits cell proliferation. Curr Biol. 2008;18:311-21
10. Zhao B, Li L, Lei Q, Guan KL. The Hippo-YAP pathway in organ size control and tumorigenesis: an updated version. Genes Dev. 2010;24:862-74
11. Zhao B, Wei X, Li W, Udan RS, Yang Q, Kim J. et al. Inactivation of YAP oncoprotein by the Hippo pathway is involved in cell contact inhibition and tissue growth control. Genes Dev. 2007;21:2747-61
12. Zhao B, Li L, Tumaneng K, Wang CY, Guan KL. A coordinated phosphorylation by Lats and CK1 regulates YAP stability through SCF(beta-TRCP). Genes Dev. 2010;24:72-85
13. Moon S, Kim W, Kim S, Kim Y, Song Y, Bilousov O. et al. Phosphorylation by NLK inhibits YAP-14-3-3-interactions and induces its nuclear localization. EMBO Rep. 2017;18:61-71
14. Ben Mimoun S, Mauviel A. Molecular mechanisms underlying TGF-beta/Hippo signaling crosstalks - Role of baso-apical epithelial cell polarity. International Journal of Biochemistry & Cell Biology. 2018;98:75-81
15. Qin Z, Xia W, Fisher GJ, Voorhees JJ, Quan T. YAP/TAZ regulates TGF-beta/Smad3 signaling by induction of Smad7 via AP-1 in human skin dermal fibroblasts. Cell Commun Signal. 2018;16:18
16. Levy D, Adamovich Y, Reuven N, Shaul Y. Yap1 phosphorylation by c-Abl is a critical step in selective activation of proapoptotic genes in response to DNA damage. Mol Cell. 2008;29:350-61
17. Piccolo S, Dupont S, Cordenonsi M. The Biology of Yap/Taz: Hippo Signaling and Beyond. Physiol Rev. 2014;94:1287-312
18. Hayashi S, Yokoyama H, Tamura K. Roles of Hippo signaling pathway in size control of organ regeneration. Dev Growth Differ. 2015;57:341-51
19. Pan D. Hippo signaling in organ size control. Genes Dev. 2007;21:886-97
20. Matsui Y, Nakano N, Shao D, Gao S, Luo W, Hong C. et al. Lats2 is a negative regulator of myocyte size in the heart. Circ Res. 2008;103:1309-18
21. von Gise A, Lin Z, Schlegelmilch K, Honor LB, Pan GM, Buck JN. et al. YAP1, the nuclear target of Hippo signaling, stimulates heart growth through cardiomyocyte proliferation but not hypertrophy. Proceedings of the National Academy of Sciences of the United States of America. 2012;109:2394-9
22. Del Re DP, Matsuda T, Zhai P, Gao S, Clark GJ, Van Der Weyden L. et al. Proapoptotic Rassf1A/Mst1 signaling in cardiac fibroblasts is protective against pressure overload in mice. J Clin Invest. 2010;120:3555-67
23. Nakamura M, Zhai P, Del Re DP, Maejima Y, Sadoshima J. Mst1-mediated phosphorylation of Bcl-xL is required for myocardial reperfusion injury. JCI Insight. 2016 1
24. Xin M, Kim Y, Sutherland LB, Murakami M, Qi X, McAnally J. et al. Hippo pathway effector Yap promotes cardiac regeneration. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:13839-44
25. Lin Z, Zhou P, von Gise A, Gu F, Ma Q, Chen J. et al. Pi3kcb links Hippo-YAP and PI3K-AKT signaling pathways to promote cardiomyocyte proliferation and survival. Circ Res. 2015;116:35-45
26. Bi L, Okabe I, Bernard DJ, Nussbaum RL. Early embryonic lethality in mice deficient in the p110beta catalytic subunit of PI 3-kinase. Mamm Genome. 2002;13:169-72
27. Xiao Y, Hill MC, Zhang M, Martin TJ, Morikawa Y, Wang S. et al. Hippo Signaling Plays an Essential Role in Cell State Transitions during Cardiac Fibroblast Development. Developmental Cell. 2018;45:153 -+
28. Billings SE, Pierzchalski K, Tjaden NEB, Pang XY, Trainor PA, Kane MA. et al. The retinaldehyde reductase DHRS3 is essential for preventing the formation of excess retinoic acid during embryonic development. Faseb Journal. 2013;27:4877-89
29. Hsu H-T, Estaras C, Huang L, Jones KA. Specifying the Anterior Primitive Streak by Modulating YAP1 Levels in Human Pluripotent Stem Cells. Stem Cell Reports. 2018;11:1357-64
30. Foulquier S, Daskalopoulos EP, Lluri G, Hermans KCM, Deb A, Blankesteijn WM. WNT Signaling in Cardiac and Vascular Disease. Pharmacol Rev. 2018;70:68-141
31. Nusse R, Clevers H. Wnt/beta-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell. 2017;169:985-99
32. Cong F, Schweizer L, Varmus H. Casein kinase Iepsilon modulates the signaling specificities of dishevelled. Mol Cell Biol. 2004;24:2000-11
33. Mlodzik M. The Dishevelled Protein Family: Still Rather a Mystery After Over 20 Years of Molecular Studies. In: Wassarman PM, editor. Essays on Developmental Biology, Pt B. 2016 p. 75-+
34. Xu F, Wang Y-l, Chang J-j, Du S-c, Diao L, Jiang N. et al. Mammalian sterile 20-like kinase 1/2 inhibits the Wnt/beta-catenin signalling pathway by directly binding casein kinase 1 epsilon. Biochemical Journal. 2014;458:159-69
35. Varelas X, Miller BW, Sopko R, Song S, Gregorieff A, Fellouse FA. et al. The Hippo pathway regulates Wnt/beta-catenin signaling. Dev Cell. 2010;18:579-91
36. Akladios B, Mendoza-Reinoso V, Samuel MS, Hardeman EC, Khosrotehrani K, Key B. et al. Epidermal YAP2-5SA-Delta C Drives beta-Catenin Activation to Promote Keratinocyte Proliferation in Mouse Skin In Vivo. Journal of Investigative Dermatology. 2017;137:716-26
37. Xin M, Kim Y, Sutherland LB, Qi X, McAnally J, Schwartz RJ. et al. Regulation of insulin-like growth factor signaling by Yap governs cardiomyocyte proliferation and embryonic heart size. Sci Signal. 2011;4:ra70
38. Klaus A, Saga Y, Taketo MM, Tzahor E, Birchmeier W. Distinct roles of Wnt/beta-catenin and Bmp signaling during early cardiogenesis. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:18531-6
39. Khan K, Makhoul G, Yu B, Schwertani A, Cecere R. The cytoprotective impact of yes-associated protein 1 after ischemia-reperfusion injury in AC16 human cardiomyocytes. Experimental Biology and Medicine. 2019;244:802-12
40. Volckaert T, Yuan T, Yuan J, Boateng E, Hopkins S, Zhang J-S. et al. Hippo signaling promotes lung epithelial lineage commitment by curbing Fgf10 and beta-catenin signaling. Development. 2019 146
41. Azzolin L, Panciera T, Soligo S, Enzo E, Bicciato S, Dupont S. et al. YAP/TAZ Incorporation in the beta-Catenin Destruction Complex Orchestrates the Wnt Response. Cell. 2014;158:157-70
42. Gargini R, Escoll M, Garcia E, Garcia-Escudero R, Wandosell F, Maria Anton I. WIP Drives Tumor Progression through YAP/TAZ-Dependent Autonomous Cell Growth. Cell Reports. 2016;17:1962-77
43. Imajo M, Miyatake K, Iimura A, Miyamoto A, Nishida E. A molecular mechanism that links Hippo signalling to the inhibition of Wnt/beta-catenin signalling. EMBO J. 2012;31:1109-22
44. Rammensee S, Kang MS, Georgiou K, Kumar S, Schaffer DV. Dynamics of Mechanosensitive Neural Stem Cell Differentiation. Stem Cells. 2017;35:497-506
45. Lim SK, Lu SY, Kang S-A, Tan HJ, Li Z, Wee ZNA. et al. Wnt Signaling Promotes Breast Cancer by Blocking ITCH-Mediated Degradation of YAP/TAZ Transcriptional Coactivator WBP2. Cancer Research. 2016;76:6278-89
46. Tang C, Takahashi-Kanemitsu A, Kikuchi I, Ben C, Hatakeyama M. Transcriptional Co-activator Functions of YAP andTAZ Are Inversely Regulated by Tyrosine Phosphorylation Status of Parafibromin. iScience. 2018;1:1-15
47. Tao H, Zhu M, Lau K, Whitley OKW, Samani M, Xiao X. et al. Oscillatory cortical forces promote three dimensional cell intercalations that shape the murine mandibular arch. Nature Communications. 2019 10
48. Feng Y, Liang Y, Zhu X, Wang M, Gui Y, Lu Q. et al. The signaling protein Wnt5a promotes TGF1-mediated macrophage polarization and kidney fibrosis by inducing the transcriptional regulators Yap/Taz. Journal of Biological Chemistry. 2018;293:19290-302
49. Park HW, Kim YC, Yu B, Moroishi T, Mo J-S, Plouffe SW. et al. Alternative Wnt Signaling Activates YAP/TAZ. Cell. 2015;162:780-94
50. Wang C, Han X, Zhou Z, Uyunbilig B, Huang X, Li R. et al. Wnt3a Activates the WNT-YAP/TAZ Pathway to Sustain CDX2 Expression in Bovine Trophoblast Stem Cells. DNA and Cell Biology. 2019;38:410-22
51. Bejoy J, Song L, Zhou Y, Li Y. Wnt/Yes-Associated Protein Interactions During Neural Tissue Patterning of Human Induced Pluripotent Stem Cells. Tissue Engineering Part A. 2018;24:546-58
52. Xiong J, Almeida M, O'Brien CA. The YAP/TAZ transcriptional co-activators have opposing effects at different stages of osteoblast differentiation. Bone. 2018;112:1-9
53. Katagiri T, Watabe T. Bone Morphogenetic Proteins. Cold Spring Harb Perspect Biol. 2016 8
54. Gomez-Puerto MC, Iyengar PV, Garcia de Vinuesa A, Ten Dijke P, Sanchez-Duffhues G. Bone morphogenetic protein receptor signal transduction in human disease. J Pathol. 2019;247:9-20
55. Huang J, Reilein A, Kalderon D. Yorkie and Hedgehog independently restrict BMP production in escort cells to permit germline differentiation in the Drosophila ovary. Development. 2017;144:2584-94
56. Karystinou A, Roelofs AJ, Neve A, Cantatore FP, Wackerhage H, De Bari C. Yes-associated protein (YAP) is a negative regulator of chondrogenesis in mesenchymal stem cells. Arthritis Research & Therapy. 2015 17
57. Ying QL, Nichols J, Chambers I, Smith A. BMP induction of Id proteins suppresses differentiation and sustains embryonic stem cell self-renewal in collaboration with STAT3. Cell. 2003;115:281-92
58. Neto F, Klaus-Bergmann A, Ong YT, Alt S, Vion A-C, Szymborska A. et al. YAP and TAZ regulate adherens junction dynamics and endothelial cell distribution during vascular development. Elife. 2018 7
59. Zeng H, Zhang Y, Yi Q, Wu Y, Wan R, Tang L. CRIM1, a newfound cancer-related player, regulates the adhesion and migration of lung cancer cells. Growth Factors. 2015;33:384-92
60. Fukui H, Miyazaki T, Chow RW-Y, Ishikawa H, Nakajima H, Vermot J. et al. Hippo signaling determines the number of venous pole cells that originate from the anterior lateral plate mesoderm in zebrafish. Elife. 2018 7
61. Bae JS, Jeon Y, Kim SM, Jang JY, Park MK, Kim I-H. et al. Depletion of MOB1A/B causes intestinal epithelial degeneration by suppressing Wnt activity and activating BMP/TGF-beta signaling. Cell Death & Disease. 2018 9
62. Zeng H, Ortiz A, Shen P-F, Cheng C-J, Lee Y-C, Yu G. et al. Angiomotin regulates prostate cancer cell proliferation by signaling through the Hippo-YAP pathway. Oncotarget. 2017;8:10145-60
63. Uemura M, Nagasawa A, Terai K. Yap/Taz transcriptional activity in endothelial cells promotes intramembranous ossification via the BMP pathway. Scientific Reports. 2016 6
64. Kiyono M, Shibuya M. Bone morphogenetic protein 4 mediates apoptosis of capillary endothelial cells during rat pupillary membrane regression. Mol Cell Biol. 2003;23:4627-36
65. Suzuki Y, Montagne K, Nishihara A, Watabe T, Miyazono K. BMPs promote proliferation and migration of endothelial cells via stimulation of VEGF-A/VEGFR2 and angiopoietin-1/Tie2 signalling. J Biochem. 2008;143:199-206
66. Valdimarsdottir G, Goumans MJ, Rosendahl A, Brugman M, Itoh S, Lebrin F. et al. Stimulation of Id1 expression by bone morphogenetic protein is sufficient and necessary for bone morphogenetic protein-induced activation of endothelial cells. Circulation. 2002;106:2263-70
67. Ruiz-Llorente L, Gallardo-Vara E, Rossi E, Smadja DM, Botella LM, Bernabeu C. Endoglin and alk1 as therapeutic targets for hereditary hemorrhagic telangiectasia. Expert Opin Ther Targets. 2017;21:933-47
68. Wooderchak-Donahue WL, McDonald J, O'Fallon B, Upton PD, Li W, Roman BL. et al. BMP9 mutations cause a vascular-anomaly syndrome with phenotypic overlap with hereditary hemorrhagic telangiectasia. Am J Hum Genet. 2013;93:530-7
69. Young K, Tweedie E, Conley B, Ames J, FitzSimons M, Brooks P. et al. BMP9 Crosstalk with the Hippo Pathway Regulates Endothelial Cell Matricellular and Chemokine Responses. Plos One. 2015 10
70. Varelas X, Samavarchi-Tehrani P, Narimatsu M, Weiss A, Cockburn K, Larsen BG. et al. The Crumbs complex couples cell density sensing to Hippo-dependent control of the TGF-beta-SMAD pathway. Dev Cell. 2010;19:831-44
71. Kumar D, Nitzan E, Kalcheim C. YAP promotes neural crest emigration through interactions with BMP and Wnt activities. Cell Communication and Signaling. 2019 17
72. Wang W, Qiao Y, Li Z. New Insights into Modes of GPCR Activation. Trends Pharmacol Sci. 2018;39:367-86
73. Yasuda D, Kobayashi D, Akahoshi N, Ohto-Nakanishi T, Yoshioka K, Takuwa Y. et al. Lysophosphatidic acid-induced YAP/TAZ activation promotes developmental angiogenesis by repressing Notch ligand Dll4. The Journal of clinical investigation. 2019 130
74. Shen Y, Zhao S, Wang S, Pan X, Zhang Y, Xu J. et al. S1P/S1PR3 axis promotes aerobic glycolysis by YAP/c-MYC/PGAM1 axis in osteosarcoma. Ebiomedicine. 2019;40:210-23
75. Feng X, Arang N, Rigiracciolo DC, Lee JS, Yeerna H, Wang Z. et al. A Platform of Synthetic Lethal Gene Interaction Networks Reveals that the GNAQ Uveal Melanoma Oncogene Controls the Hippo Pathway through FAK. Cancer Cell. 2019;35:457 -+
76. An J, Li G, Zhang J, Zhou H, Jiang J, Wang X. et al. GNAS knockdown suppresses osteogenic differentiation of mesenchymal stem cells via activation of Hippo signaling pathway. Journal of cellular physiology. 2019
77. Yui OM, Benitez JA, Plouffe SW, Ryback D, Klein A, Smith J. et al. YAP and MRTF-A, transcriptional co-activators of RhoA-mediated gene expression, are critical for glioblastoma tumorigenicity. Oncogene. 2018;37:5492-507
78. Yu OM, Miyamoto S, Brown JH. Myocardin-Related Transcription Factor A and Yes-Associated Protein Exert Dual Control in G Protein-Coupled Receptor- and RhoA-Mediated Transcriptional Regulation and Cell Proliferation. Molecular and Cellular Biology. 2016;36:39-49
79. Regue L, Mou F, Avruch J. G protein-coupled receptors engage the mammalian Hippo pathway through F-actin F-Actin, assembled in response to Galpha(12/13) induced RhoA-GTP, promotes dephosphorylation and activation of the YAP oncogene. Bioessays. 2013;35:430-5
80. Wang J, Hong Y, Shao S, Zhang K, Hong W. FFAR1-and FFAR4-dependent activation of Hippo pathway mediates DHA-induced apoptosis of androgen-independent prostate cancer cells. Biochemical and Biophysical Research Communications. 2018;506:590-6
81. Thirunavukkarasan M, Wang C, Rao A, Hind T, Teo YR, Siddiquee AA-M. et al. Short-chain fatty acid receptors inhibit invasive phenotypes in breast cancer cells. Plos One. 2017 12
82. Zhang K, Hu Z, Qi H, Shi Z, Chang Y, Yao Q. et al. G-protein-coupled receptors mediate omega-3 PUFAs-inhibited colorectal cancer by activating the Hippo pathway. Oncotarget. 2016;7:58315-30
83. Wang Z, Sun L, Liang S, Liu Z-c, Zhao Z-y, Yang J. et al. GPER stabilizes F-actin cytoskeleton and activates TAZ via PLC beta-PKC and Rho/ROCK-LIMK-Cofilin pathway. Biochemical and Biophysical Research Communications. 2019;516:976-82
84. Alarcon VB, Marikawa Y. ROCK and RHO Playlist for Preimplantation Development: Streaming to HIPPO Pathway and Apicobasal Polarity in the First Cell Differentiation. In: Knott J, Latham K, editors. Chromatin Regulation of Early Embryonic Lineage Specification. 2018 p. 47-68
85. Serafimidis I, Rodriguez-Aznar E, Lesche M, Yoshioka K, Takuwa Y, Dahl A. et al. Pancreas lineage allocation and specification are regulated by sphingosine-1-phosphate signalling. Plos Biology. 2017 15
86. Kemppainen K, Wentus N, Lassila T, Laiho A, Tornquist K. Sphingosylphosphorylcholine regulates the Hippo signaling pathway in a dual manner. Cellular Signalling. 2016;28:1894-903
87. Jiang LI, Collins J, Davis R, Lin KM, DeCamp D, Roach T. et al. Use of a cAMP BRET sensor to characterize a novel regulation of cAMP by the sphingosine 1-phosphate/G13 pathway. J Biol Chem. 2007;282:10576-84
88. Moroishi T, Park HW, Qin B, Chen Q, Meng Z, Plouffe SW. et al. A YAP/TAZ-induced feedback mechanism regulates Hippo pathway homeostasis. Genes Dev. 2015;29:1271-84
89. Deng Q, Jiang G, Wu Y, Li J, Liang W, Chen L. et al. GPER/Hippo-YAP signal is involved in Bisphenol S induced migration of triple negative breast cancer (TNBC) cells. Journal of Hazardous Materials. 2018;355:1-9
90. Van Kolen K, Slegers H. Integration of P2Y receptor-activated signal transduction pathways in G protein-dependent signalling networks. Purinergic Signal. 2006;2:451-69
91. Ellison GM, Vicinanza C, Smith AJ, Aquila I, Leone A, Waring CD. et al. Adult c-kit(pos) cardiac stem cells are necessary and sufficient for functional cardiac regeneration and repair. Cell. 2013;154:827-42
92. Khalafalla FG, Greene S, Khan H, Ilves K, Monsanto MM, Alvarez R Jr. et al. P2Y(2) Nucleotide Receptor Prompts Human Cardiac Progenitor Cell Activation by Modulating Hippo Signaling. Circulation Research. 2017;121:1224-36
93. Hao F, Xu Q, Zhao Y, Stevens JV, Young SH, Sinnett-Smith J. et al. Insulin Receptor and GPCR Crosstalk Stimulates YAP via PI3K and PKD in Pancreatic Cancer Cells. Molecular Cancer Research. 2017;15:929-41
94. Wang J, Sinnett-Smith J, Stevens JV, Young SH, Rozengurt E. Biphasic Regulation of Yes-associated Protein (YAP) Cellular Localization, Phosphorylation, and Activity by G Protein-coupled Receptor Agonists in Intestinal Epithelial Cells: A NOVEL ROLE FOR PROTEIN KINASE D (PKD). Journal of Biological Chemistry. 2016;291:17988-8005
95. Rao R, Salloum R, Xin M, Lu QR. The G protein G alpha(s) acts as a tumor suppressor in sonic hedgehog signaling-driven tumorigenesis. Cell Cycle. 2016;15:1325-30
Author contact
![]() Corresponding authors: Ning Hou, Key Laboratory of Molecular Target & Clinical Pharmacology, School of Pharmaceutical Sciences and the Fifth Affiliated Hospital, Guangzhou Medical University, Guangzhou 511436, China. E-mail address: houningedu.cn. Jiandong Luo, Guangzhou Institute of Cardiovascular Disease, Guangzhou Key Laboratory of Cardiovascular Disease, and the Second Affiliated Hospital, Guangzhou Medical University, Guangzhou 510260, P.R. China. E-mail address: jiandongluocom.
Corresponding authors: Ning Hou, Key Laboratory of Molecular Target & Clinical Pharmacology, School of Pharmaceutical Sciences and the Fifth Affiliated Hospital, Guangzhou Medical University, Guangzhou 511436, China. E-mail address: houningedu.cn. Jiandong Luo, Guangzhou Institute of Cardiovascular Disease, Guangzhou Key Laboratory of Cardiovascular Disease, and the Second Affiliated Hospital, Guangzhou Medical University, Guangzhou 510260, P.R. China. E-mail address: jiandongluocom.