Impact Factor ISSN: 1449-2288
- Issue 12; 2026
- Issue 11; 2026
- Issue 10; 2026
- Issue 9; 2026
- Issue 8; 2026
- Volume 22; 2026
- Past Issues
- Advance Articles
- Editorial Board
- Cover Images
- Index & Coverage
- Cover Suggestion
- Special Issues
1. Introduction
2. Gut microbiota and...
3. Microbial-derived metabolites...
4. The therapeutic strategies in...
5. Conclusion, perspective and...
Acknowledgements
References

 Global reach, higher impact
Global reach, higher impactInt J Biol Sci 2026; 22(10):5421-5443. doi:10.7150/ijbs.133969 This issue Cite
Review
Gut microbial-derived metabolites: key players in kidney disease and renal fibrosis
Wan-Ying Zhang1#, Yu-Lu Zhang1#, Qing-Qing Yu1#, Jing-Teng Zhou1, Hua Miao1,2 ![]() , Li-Min Liu1,3
, Li-Min Liu1,3 ![]() , Ying-Yong Zhao1,2,4
, Ying-Yong Zhao1,2,4 ![]()
1. School of Pharmaceutical Sciences, Zhejiang Chinese Medical University, No. 548 Binwen Road, Hangzhou, Zhejiang 310053, China.
2. School of Pharmaceutical Sciences, The First Affiliated Hospital of Zhejiang Chinese Medical University, No. 54 Youdian Road, Hangzhou 310003, China.
3. School of Medicine, Northwest University, No. 229 Taibai North Road, Xi'an, Shaanxi, 710069, China.
4. State Key Laboratory of Kidney Diseases, First Medical Center of Chinese PLA General Hospital, No. 28 Fuxing Road, Beijing 100853, China.
#Equal contribution.
Received 2026-3-5; Accepted 2026-4-30; Published 2026-5-18
Abstract

Kidney disease is a major global public health problem that affects 15-20% of adults globally. Increasing evidence shows that kidney injury results in intestinal barrier dysfunction, microbial dysbiosis, and microbial-derived metabolite disorder. Microbial-derived metabolites are recognized as multi-kingdom intermediates. The alterations of the gut microbiota lead to the reductions of short-chain fatty acids including acetate, butyrate, and propionate, while excessive accumulation of uremic toxins, including indoxyl sulfate, indole-3-acetic acid, trimethylamine-N-oxide, and p-cresyl sulphate that involved in renal disease. Various effects are moderated by regulating aryl hydrocarbon receptor, G-protein-coupled receptor 43, Toll-like receptor 4, domain-like receptor family protein 3, phosphatidylinositol-3 kinase, inhibitor of kappa B /nuclear factor kappa B and kelch-like ECH-associated protein 1/nuclear factor erythroid 2-related factor 2 pathways via gut microbiota-derived metabolites. Among them, tryptophan metabolites are considered pivotal in the communication between gut microbiota and kidneys. This review summarizes current understanding of the role of gut microbial-derived metabolites in acute kidney injury and chronic kidney disease including diabetic kidney disease, immunoglobulin A nephropathy and membranous nephropathy, explain the underlying pathophysiology of these associations and pinpoint potential targets for the future precision-based modulation of therapies. This review also explores therapeutic options, such as natural products, prebiotics, probiotics, and renal replacement therapy, for targeting gut microbiota dysbiosis and their metabolites in patients with kidney diseases to provide a more concept-driven and precise therapy strategy.
Keywords: gut microbiota, acute kidney injury, chronic kidney disease, renal fibrosis, short-chain fatty acids, natural products
1. Introduction
Kidney disease is a major global health burden that affects more than 15-20% of the worldwide population [1, 2]. Kidney disease including acute kidney injury (AKI) and chronic kidney disease (CKD) is characterized by a progressive decline in renal function accompanied by considerable morbidity and mortality [3, 4]. Current interventions for renal disease, including renal transplantation and dialysis, are costly and not curative [5-7]. Renal fibrosis, including renal tubulointerstitial fibrosis and glomerular sclerosis, is a chronic and progressive process [8]. Renal fibrosis is characterized by massive activation and proliferation of myofibroblasts, macrophages and mesangial cells that lead to the excessive deposition of extracellular matrix and tissue scarring [9-13]. Regardless of the underlying aetiology, is the ultimate common pathway for almost all types of chronic repetitive injury in the kidneys, ultimately leading to end-stage renal disease (ESRD) [9, 14]. Substantial publications have dictated that renal fibrosis is extensively related to diverse signals such as hyperactive renin-angiotensin system, high mobility group box 1 and aryl hydrocarbon receptor (AHR) [15-17], and profibrotic pathways such as inhibitor of kappa B(IƙB)/nuclear factor kappa B (NF-ƙB), transforming growth factor β1 (TGF-β1)/suppressor of mothers against decapentaplegic (Smad) and Wnt1/β-catenin [18, 19]. Moreover, emerging multi-omics (genomics, epigenetics, transcriptomics, proteomics, metabolomics, lipidomics and phenomics)-based molecular mechanisms highlighted that renal fibrosis is associated with gut microbial dysbiosis [20, 21], non-coding ribonucleic acids (ncRNAs) dysregulation [21-24] and altered levels in metabolites including tryptophan and lipid (phosphatidylcholines and bile acids) metabolisms [25-29].
Ample evidence has shown that renal fibrosis leads to intestinal epithelial cell barrier injury that increases intestinal permeability and gut microbiota dysbiosis that causes altered microbial-derived metabolites, such as indoxyl sulfate (IS), trimethylamine-N-oxide (TMAO), p-cresyl sulphate (PCS), indole-3-acetic acid (IAA), hyodeoxycholic acid and 1,5-anhydroglucitol, that correlate with renal functions [30, 31]. These metabolites leak into bloodstream through damaged intestinal epithelial cell barrier, and mediate systemic and local low-level inflammation that further aggravates kidney injury and exacerbates progressive renal fibrosis by regulating a variety of signalling pathways, such as AHR, farnesoid X receptor, glucagon-like peptide-1 receptor (GLP-1), and transmembrane G protein-coupled bile acid receptor 5 [32-34]. Recently, integrating metagenomics and metabolomics reveals that renal fibrosis is related to gut microbial dysbiosis that leads to the aberrant microbial-derived metabolites [35, 36]. Oxidative stress and inflammation play a central role in the progression and outcome of renal disease [37]. Studies have revealed that CKD results in intestinal inflammation and impaired epithelial barrier function, which leads to an accelerated systemic translocation of bacteria-derived uremic toxins [38]. Moreover, the aberrant gut microbiota in CKD patients results in metabolite disorder and ultimately exacerbates clinical outcomes [36, 39-41]. Therefore, modulating gut-metabolite-kidney axis could be an effective approach to disease intervention.
This review summarizes the recent insights into the roles of gut microbiota-derived metabolites, such as short-chain fatty acids (SCFAs), microbial-derived tryptophan metabolites including IS, IAA, indole-3-propionic acid (IPA) and indole-3-aldehyde (IAld), TMAO, PCS and polyamines in health and kidney disease. The focus is on the regulatory functions of microbial metabolites in kidney disease, and their potential role in facilitating communication between gut and kidneys is explored. We also discuss current therapeutic strategies targeting the modulation of gut microbiota-derived metabolites through dialysis (hemodialysis, peritoneal dialysis and colonic dialysis), kidney transplantation (KT), natural product interventions, microecological therapies (probiotics, prebiotics and synbiotics), and dietary interventions in the management of kidney disease.
2. Gut microbiota and microbial-derived metabolites in health
2.1. Gut microbiota in health
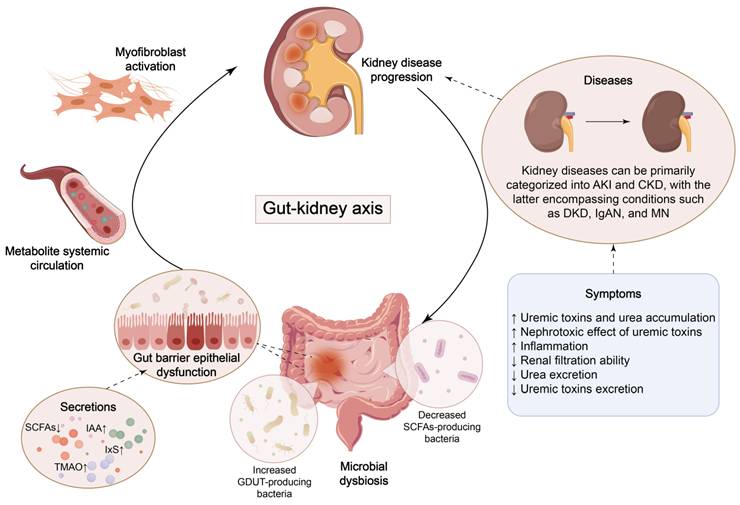
Over the past decade, research has demonstrated that the extensive community of micro-organisms residing in the gut, known as the gut microbiota, is closely associated with human health and disease, partly due to its impact on systemic immune responses [42, 43]. Gut microbiota can synthesize multiple vitamins and essential amino acids necessary for human development and growth, promote intestinal peristalsis, and maintain intestinal function [44]. Four dominant phyla, Firmicutes, Bacteroidetes, Actinobacteria, and Verrucomicrobia, account for 99% of healthy human gut bacteria, with Firmicutes and Bacteroidetes dominating at a combined proportion of around 90% [45-47]. Under normal circumstances, the gut microbiota maintains a symbiotic relationship with the host, which contribute to the therapy of metabolic dysfunction in humans, influencing intestinal immunity, tolerance, and sensitivity to inflammation [48]. The theory of the gut-kidney axis has arisen collectively, highlighting the bidirectional communication between renal function and gut microbiota (Figure 1).
Crosstalk between kidney disease and microbial dysbiosis. Dysbiosis of gut bacteria leads to the accumulation of uremic toxins and impairment of the gut barrier, which triggers renal inflammation and fibrosis. The progression of kidney disease, in turn, exacerbates gut dysfunction.
2.2. Gut microbiota and microbial-derived metabolites in health
2.2.1. SCFAs
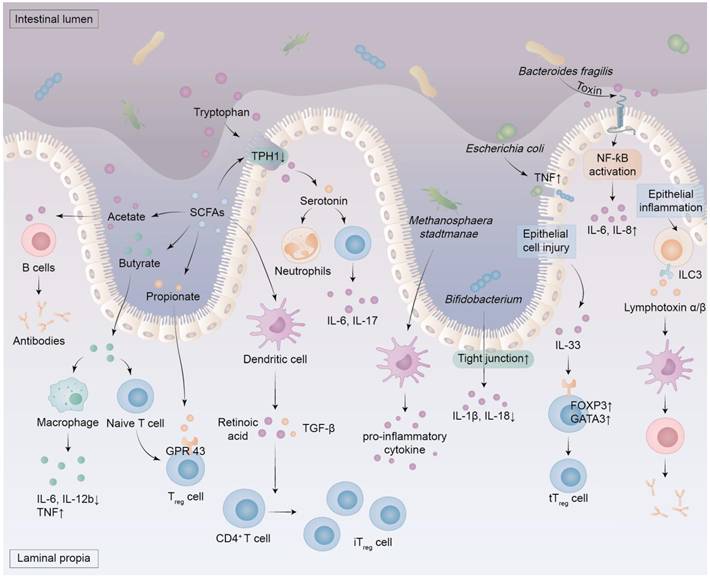
Most colonic bacteria are strict anaerobes that ferment indigestible complex carbohydrates into SCFAs, such as acetate, butyrate, and propionate (Figure 2) [49]. Their proportions vary according to diet, with propionate generated by some species from Firmicutes and Bacteroidetes accounting for 10-20%, acetate generated by several bacterial groups accounting for 50-70%, and butyrate generated by a few Clostridia species constituting the majority of the remainder [50]. Notably, Mitsuokella jalaludinii is an effective phytate degrader that can synergize with Anaerostipes rhamnosivorans to generate propionate [51]. SCFAs play an important role in human health and disease [52]. For instance, SCFAs produced by gut microbiota act as vital signalling molecules and metabolic substrates, impacting the host's metabolism, immune system, and gut-brain communication [52, 53]. The SCFAs regulate extra-intestinal immunity directly and indirectly, and are associated with various disorders, including autoimmunity, infections, and intestinal inflammation [49, 52]. The molecular mechanisms by which SCFAs regulate physiological processes involve the deorphanization of free fatty acid receptors 2 and 3 [54]. They regulate inflammation and barrier function via G protein-coupled receptors and histone deacetylase inhibition, with therapeutic potential in chronic inflammatory conditions [49]. The SCFAs attenuate intestinal inflammation by striking a balance of regulatory T (Treg)/T helper 17 cells, while also lowering the levels of pro-inflammatory cytokines such as interleukin-6 (IL-6), IL-1β, and tumor necrosis factor-alpha (TNF-α) [55]. The anti-inflammatory role of butyrate stems from direct modulation of phagocytes, intestinal epithelial cells, plasma cells, B cells, effector and Treg cells (Figure 2) [49]. Due to its anti-inflammatory properties, high levels of butyrate are associated with improved intestinal immunity and intestinal barrier integrity, making it both nephroprotective and enteroprotective [56]. In addition, propionate and butyrate serve as unique epigenetic regulatory elements linking diet, metabolism, and gene expression, and influence the proliferation of both normal cells and colorectal cancer cells [53]. Overall, SCFAs serve as key mediators of the interaction between the host and the microbiota, and targeted supplementation with SCFAs may represent a promising therapeutic strategy for disease treatment [57].
The role of metabolites and components produced by the dysbiosis of intestinal microbiota. Dysbiosis of the intestinal microbiota results in an elevation of pathogen levels and intestinal uremic toxins, which subsequently compromises the integrity of the intestinal barrier and instigates oxidative stress and inflammation within the intestinal mucosa. This process activates immune cells that secrete pro-inflammatory factors. Notably, certain components of this microbiota exacerbate the pro-inflammatory response by engaging the NF-ƙB signaling pathway. In cases of intestinal dysbiosis, there is a reduction in the production of SCFAs derived from dietary fiber fermentation by specific bacteria. This decline diminishes tryptophan hydroxylase 1 activity in enterochromaffin cells located within the gastrointestinal epithelium, resulting in decreased serotonin levels alongside increased tryptophan concentrations. Furthermore, SCFAs can activate dendritic cells to release TGF-β and retinoic acid, thereby facilitating the differentiation of CD4+ T cells into iTreg cells. SCFAs include acetate, propionate, and butyrate, among others. Butyrate enhances macrophage polarization and anti-inflammatory responses by inhibiting TNF production while down-regulating IL-6 and IL-12b expression. It also promotes naive T cell differentiation into Treg cells. Propionate stimulates GPR43 on Treg cells, thereby augmenting their proliferation, whereas acetate fosters antibody synthesis in B cells. Inflammation of the epithelium induces ILC3 to produce lymphotoxin α and β, which leads to the activation of dendritic cells and helps B cells produce antibodies. Damage to epithelial cells prompts IL-33 release which increases GATA3 and FOXP3 expression in T cells, ultimately promoting both proliferation and maintenance of tTreg cells. FOXP3, forkhead box protein 3; GATA3, GATA binding protein 3; ILC3, group 3 innate lymphoid cell; iTreg cells, induced regulatory T cells; TPH 1, tryptophan hydroxylase 1; tTreg cells, thymic regulatory T cells.
2.2.2. Microbial-derived tryptophan metabolites: AHR ligands
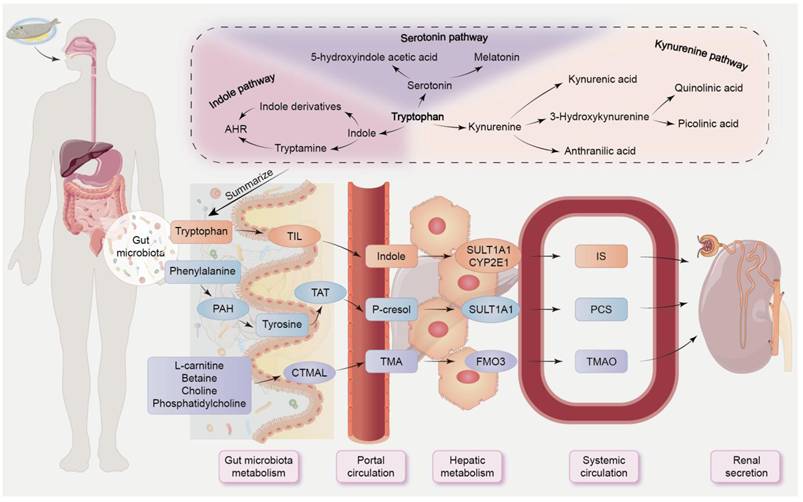
AHR, as a ligand-dependent transcription factor, regulates intricate transcriptional processes in diverse cell types and exerts primary anti-inflammatory and protective effects in the gut via innate lymphoid cells and IL-22 [50, 58]. Many tryptophan metabolites, such as IS, IAA, IPA and IAld produced by the gut microbiota, can act as AHR ligands, inducing AHR activation [58]. Our previous research revealed that the plasma tryptophan level was negatively correlated with Intestinimonas, Blautia, and Oscillibacter, but positively correlated with Lactobacillales, Pseudomonas, Turicibacter, and Clostridium IV in rats with unilateral ureteral obstruction [48]. Tryptophan can be catabolized through three different pathways: serotonin/melatonin pathway (≈1%-2%), indole pathway (≈5%), and kynurenine pathway (≈95%) (Figure 3) [32, 59]. A small portion of dietary tryptophan is degraded in gut by bacterial tryptophan indole lyase to form indole, which is subsequently absorbed into portal circulation, and then converted to indoxyl, which is then transformed into indoxyl acetate and IS for excretion through the kidneys (Figure 3) [60, 61]. Indole stimulates the release of GLP-1, while indole derivatives, including IAA, IAld, indole-3-acetaldehyde and indoleacrylic acid, can act as AHR agonists [50]. Tryptophan supplementation reshapes gut microbiota, alleviates tissue damage, and enhances integrity of both intestinal epithelial and gut vascular barriers [62]. Indole-3-lactic acid treatment protects the intestinal vascular barrier by downregulating claudin-2 via AHR/nuclear factor erythroid 2-related factor 2 (Nrf2)/signal transducer and activator of transcription 3-mediated pathways in mice induced by intestinal ischemia-reperfusion (I/R) [62]. The latest study revealed that renal fibrosis is mediated by Wnt/β-catenin pathway via gut microbial-mediated tryptophan metabolism-driven AHR signaling [63].
The absorption and excretion pathways of gut-derived uremic toxins. Some dietary breakdown products are excreted in faeces, while others enter portal circulation via efflux and influx transporters on colonocytes. The end products are then eliminated by healthy kidneys. Tryptophan can be catabolized by three different metabolic pathways: the serotonin pathway, the indole pathway, and the kynurenine pathway. In the indole pathway, tryptophan is converted to indole by TIL and subsequently absorbed into the portal circulation. Subsequently, it is metabolized to indoxyl sulfate by sulfotransferase 1A1 and cytochrome P450 2E1. Phenylalanine undergoes hydroxylation to form tyrosine through the action of intestinal bacteria utilizing PAH, with tyrosine further being metabolized to p-cresol via tyrosine aminotransferase. These intermediate metabolites are subsequently converted to p-cresyl sulfate by sulfotransferase 1A1. Choline and other dietary molecules are degraded into TMA through carnitine trimethylamine lyase, followed by oxidation to TMAO mediated by flavin-containing monooxygenase 3. CTMAL, carnitine trimethylamine lyase; CYP2E1, cytochrome P450 2E1; PAH, phenylalanine hydroxylase; TIL, tryptophan indole lyase; FMO3, flavin-containing monooxygenase 3; SULT1A1, sulfotransferase 1A1; TAT, tyrosine aminotransferase.
2.2.3. TMAO
TMAO is produced through hepatic processing of trimethylamine (TMA), which is derived from gut bacteria and is metabolised from certain dietary nutrients, such as choline, L-carnitine, betaine and phosphatidylcholine (Figure 3) [64, 65]. Emergencia timonensis can catalyze the conversion of carnitine to the intermediate γ-butyrobetaine, which then converts to TMA, a precursor of TMAO [66]. Plasma γ-butyrobetaine levels are significantly linked to incident cardiovascular disease (CVD) risk in a clinical cohort [66]. Increasing evidence indicates that targeting uremic toxin-induced pathways or uremic toxins, including TMAO and tryptophan metabolites, may reduce the risk of CVD patients with CKD [67]. TMAO, produced by liver and aorta, enhances trained immunity by increasing inflammation through endoplasmic reticulum stress, mitochondrial reactive oxygen species (ROS), and glycolysis pathways [68].
2.2.4. PCS
PCS originates from the microbial metabolite p-cresol, which is derived from phenylalanine and tyrosine. Once absorbed into the portal circulation, p-cresol is converted by hepatic enzymes into harmful uremic toxins. The end-products of microbial protein fermentation are filtered out by tubular secretion and glomerular filtration (Figure 3) [69]. Beyond renal excretion, the transport of PCS and IS, along with their microbiota-derived precursors p-cresol and indole, across key physiological barriers, including the blood-brain barrier, intestinal barrier, and renal proximal tubule, is also crucial for their elimination and distribution. Due to their high protein binding, PCS and IS are difficult to remove effectively by dialysis [70]. The p-cresol has antioxidant and antimicrobial properties, but has also been linked to kidney disease, enteric pathogens, and autism [71]. Molecular mechanisms of PCS include induction of apoptotic pathways, direct damage of cell membranes, activation of c-Jun N-terminal protein kinase and p38-mitogen-activated protein kinase, and activation of nicotinamide adenine dinucleotide phosphate oxidase 4 (NOX4) leading to ROS formation [72].
2.2.5. Polyamines
The gut microbiota produces polyamines, including cadaverine, spermine, spermidine, and putrescine, which are essential for normal bacterial cell growth and multiplication [73]. Polyamines exert anti-inflammatory effects by maintaining mitochondrial function and promoting autophagy, thereby conferring organ-protective and cytoprotective effects [74]. Putrescine, which is derived from commensal bacteria, increases anti-inflammatory macrophages in the colon. As a substrate for symbiotic metabolism, putrescine is further absorbed and metabolized by the host, thereby contributing to the maintenance of intestinal mucosal homeostasis [75]. Due to their polycationic nature, polyamines readily bind to anions in cells. Instead of binding to cytoplasmic proteins, intracellular polyamines primarily form polyamine-RNA complexes, stabilizing them [73]. Small RNAs in extracellular vesicles derived from Lactobacillus murinus downregulate host polyamine metabolism by targeting enzymes involved in polyamine metabolism. Additionally, Lactobacillus murinus impeded the recovery of dextran sodium sulfate-induced colitis in mice by decreasing polyamine levels [76]. Loss of spermine oxidase (SMOX), which produces spermidine, has been linked to increased α-defensin expression, exacerbated colitis, and microbiota dysbiosis by increasing abundance of Deferribacteres and Proteobacteria and reducing Prevotella abundance. Supplementing with spermine reverses these phenotypes, suggesting that it could be used as an additional treatment for colitis [77].
3. Microbial-derived metabolites as intermediates in gut-kidney communication
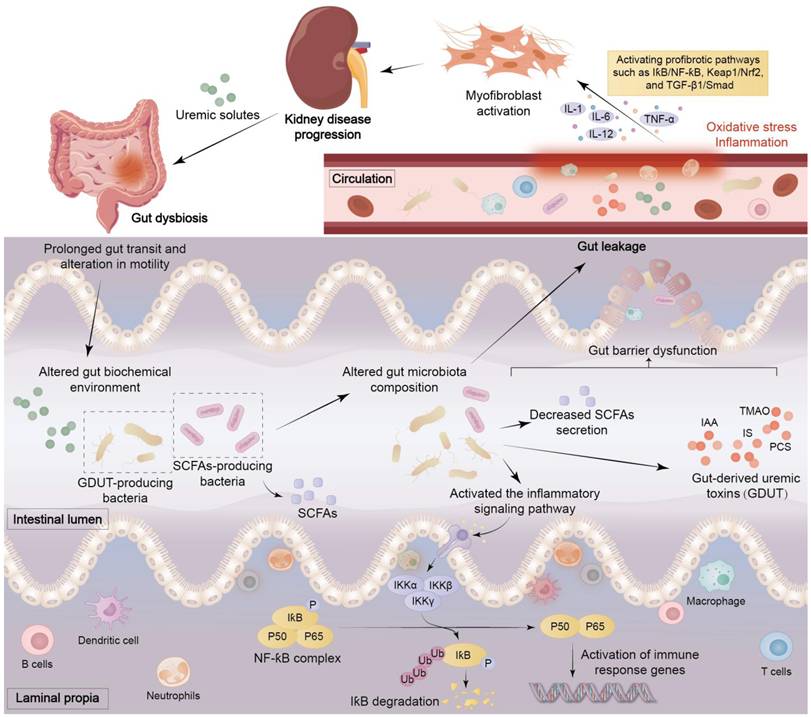
Renal disease can lead to dysbiosis of the intestinal microbiota, which can destroy the gut mucosal barrier and permit live bacteria or their metabolites to enter the bloodstream. The translocation and leakage of these metabolites across the intestinal barrier into the circulation trigger inflammation and oxidative stress, which ultimately lead to vascular and local tissue dysfunction [20, 32, 35]. Renal disease leads to an increase in levels of uremic toxins originating from the colon. The features of the uremic microenvironment are intestinal microbial dysbiosis, intestinal epithelial cell barrier impairment, and immune cell dysfunction (Figure 4) [38]. The uremic microenvironment leads to changes in the composition of microorganisms, infiltration of inflammatory cells, overproduction of immune complexes and antibodies, and abnormal activation of immune cells, which cause oxidative stress and inflammation that further exacerbate damage to renal parenchyma (Figures 2, 4) [48].
The mechanisms of interaction between intestinal and kidney diseases. Uremic solutes generated by kidney diseases can trigger gut bacteria dysbiosis, leading to prolonged intestinal transit time and altered peristalsis. The intestinal biochemical environment is then subject to alteration, affecting the composition of the intestinal microbiota, reducing the secretion of SCFAs, producing gut-derived uremic toxins (GDUT), and abnormally activating immune cells and inflammatory signalling pathways such as NF-ƙB. These factors have been demonstrated to induce intestinal barrier dysfunction, thereby precipitating leaky gut and bacterial translocation. Once uremic toxins enter the circulatory system, they trigger oxidative stress and inflammatory responses, activating profibrotic pathways such as IƙB/NF-ƙB, Keap1/Nrf2, and TGF-β1/Smad, prompting immune cells to secrete IL-1, IL-12, IL-6, and TNF-α, ultimately inducing renal fibrosis. This creates a vicious cycle that exacerbates kidney damage.
3.1. Intestinal epithelial cell barrier dysfunction in renal disease
AKI is a complex clinical syndrome that affects approximately 13.3 million people worldwide each year [78, 79]. AKI is characterized by an abrupt drop in glomerular filtration rate (GFR), a reduction in urine output, a rise in serum electrolytes, urea and creatinine, which results in a rapid loss of kidney function and augments the risk of CKD [80, 81]. AKI provokes intestinal inflammation and increased gut permeability [20]. In pathophysiological conditions, proinflammatory cytokines, antigens and pathogens such as Clostridium perfringens, Escherichia coli, and Vibrio cholerae, cause intestinal epithelial cell barrier impairment [82]. During AKI sepsis, the mucosal membrane barrier is disrupted, intensifying systemic inflammation and promoting aggravation of AKI [83]. This suggests a potential link between intestines and kidneys (Figure 1). In severe acute pancreatitis, TNF-α elevates gut permeability, induces bacterial translocation from epithelium, ulteriorly irritates massive release of inflammatory cytokines, which probably exacerbate AKI [84].
CKD is a major global health problem that affects more than 10-15% of the adult population [85, 86]. CKD is a progressive disease characterized by changes in the function and structure of the kidneys and is generally defined as persistent (>3 months) renal functional loss, high albuminuria or low GFR, eventually resulting in ESRD [87, 88]. CKD patients show increased intestinal permeability and protective mucus layer degradation [89]. Intestinal barrier dysfunction facilitates persistent leakage of metabolites through intestinal wall into systemic circulation and hepatic portal system, activating Toll-like receptors alongside inflammasome-mediated responses in innate immune cells as well as vascular endothelial cells across regional, local, and systemic sites [90]. This triggers proinflammatory cytokine release and shedding of monocyte receptors, such as lipopolysaccharide (LPS) receptor, driving chronic inflammation typical of CKD [90]. An analysis involving 48 children with CKD revealed stage-dependent increases in serum TNF-α, soluble CD14 and tryptophan metabolites alongside diminished production of SCFAs, suggesting gut barrier dysfunction, inflammation, and endotoxemia [91]. Dodd et al. found that lower intestinal IPA impairs intestinal barrier integrity, elevating permeability and activating inflammation [92]. As mentioned above, inflammatory factors and uremic toxins produced by gut microbiota during renal disease can lead to intestinal epithelial barrier dysfunction. Theoretically, gut microbial dysbiosis can compromise the health of this barrier.
3.2. Gut microbial dysbiosis in renal disease
3.2.1 Gut microbial dysbiosis in AKI
Specific members within the intestinal microbial community may ameliorate or, conversely, exacerbate ischemic renal injury [93]. I/R injury constitutes one of the primary causes of AKI [94]. An increase in Enterobacteriaceae and a decrease in Ruminococcaceae and Lactobacilli were revealed to be hallmarks of I/R injury-induced dysbiosis and related to decreased levels of SCFAs, leaky gut, and intestinal inflammation [93]. In LPS-induced AKI mouse models, pretreatment with the probiotic Akkermansia muciniphila substantially attenuated kidney injury and reduced tubular necrosis and apoptosis. This protective effect was accompanied by marked shifts in intestinal microbial ecology and preserved gut barrier integrity, demonstrating that modulating gut dysbiosis can ameliorate AKI outcomes [95]. Therefore, the changes in the gut microbiota occur in the progression of AKI.
It is worth noting that AKI progression also affects the composition of the gut microbiota. Clinically, AKI-induced gut microbiota dysbiosis includes disturbances in Bifidobacterium, Bacteroidetes, Salmonella, Staphylococcus, and Faecalibacterium [93]. Another study indicated that the abundance of Bifidobacterium was decreased while the abundance of Ruminococcus and Clostridium was increased after I/R injury [96]. In renal I/R injury models, AKI was found to downregulate enzymes in the intestinal transsulfuration pathway, reduce glutathione levels, impair tight junctions, and disrupt gut microbiota composition [97]. In summary, kidney damage can alter the structure and composition of the gut microbiota.
3.2.2 Gut microbial dysbiosis in CKD
The gut microbiota contributes to the progression of CKD by producing uremic toxins and inducing systemic inflammation [98]. The accumulation of bacterial species that produce uremic toxins over time was associated with worsening renal function. The abundance of these bacterial species was significantly higher in patients with severe CKD than in those with moderate CKD [99]. Additionally, Bacteroides fragilis has been shown to mitigate renal fibrosis by increasing 1,5-anhydroglucitol concentrations and decreasing LPS levels [34]. A prominent characteristic of diabetic kidney disease (DKD) is a lower ratio of Firmicutes (gram-positive) to Bacteroidetes (gram-negative), which positively correlates with plasma glucose levels [100]. Notably, the relative abundance of g_Prevotella_9 can accurately predict individuals with diabetes [69]. In diabetic mice, the presence of gut-derived Klebsiella oxytoca in the kidneys and circulation, alongside elevated IL-17, was found to induce the expression of kidney injury molecule-1, thereby promoting DKD progression [101]. The significant expansion of the Escherichia-Shigella genus may serve as therapeutic targets and promising diagnostic biomarkers for immunoglobulin A nephropathy (IgAN) [102]. Notably, the abundance of Bifidobacterium was decreased in both IgAN patients and mice, and the proportion of Bifidobacterium was negatively correlated with hematuria and proteinuria levels, suggesting that reduced Bifidobacterium abundance might be associated with IgAN severity [103]. A negative correlation was revealed between the abundance of Escherichia-Shigella and proteinuria levels, while the abundance of Bacteroides and Klebsiella was positively correlated with membranous nephropathy (MN) stage [39]. Moreover, the increase of pathogenic bacteria and the reduction of Roseburia and certain species of Faecalibacterium that can ferment various sugars into SCFAs may cause or exacerbate primary MN [104].
The uremic environment caused by CKD, characterized by elevated blood urea levels, metabolic acidosis, and systemic inflammation, can alter the composition of the gut microbiota. CKD patients exhibited reduced microbiota diversity, altered community structures, disrupted potential functions and microbial composition [30, 105]. Mollicutes and Tenericutes were enriched in CKD patients at tages 1-2, Parasutterella was enriched in CKD patients at stages 3-4, and Blautia and Akkermansia were enriched in CKD patients at stage 5 [106]. Ren et al. found that the abundances of Akkermansia and Thalassospira were increased with CKD progression [106]. Wu et al. discovered that a core CKD-related microbiota consists of 7 bacteria genera and 2 bacteria species including Bacteroides eggerthii and Collinsella stercoris, which are associated with the stages of CKD [107]. The discriminative species for DKD patients with microalbuminuria include Acinetobacter johnsonii and Escherichia coli, whereas for DKD patients with macroalbuminuria, Enterococcus faecalis, Morganella morganii, and Pseudomonas_E oleovorans were identified [108]. Studies have shown that the dysbiosis observed in patients with CKD is not static, but worsens progressively as the disease advances. Furthermore, over time, the proportion of uremotoxin-producing bacteria gradually increased, while the overall abundance of microbiota decreased [99]. This suggests that declining renal function directly drives the dynamic evolution of the gut microbiota composition. In summary, renal disease and gut microbial dysbiosis create a vicious cycle that exacerbates each other and accelerates disease progression.
3.3. Microbial-derived metabolites in renal disease
3.3.1. SCFAs
Recent studies have revealed that intestinal microbiota-associated metabolites, particularly SCFAs, are closely correlated with renal disease [109]. L. casei Zhang increased nicotinamide and SCFAs levels in the serum and kidneys of mice with bilateral renal I/R, mitigating renal damage and slowing the progression to CKD [41]. Treatment with three major SCFAs (acetate, propionate, and butyrate) improved renal dysfunction in mice with I/R, associated with reduced levels of local and systemic inflammation, cell activation/infiltration, and apoptosis, increased autophagy, and oxidative cellular stress [110]. Treatment of mice with acetate-producing bacteria also resulted in improved outcomes following AKI [110].
It was found that Faecalibacterium prausnitzii, a bacterium that produces butyrate and has anti-inflammatory properties, was depleted in CKD patients during its progression to ESRD. Consequently, butyrate levels decreased and intestinal homeostasis was disrupted [111]. Butyrate partially ameliorates renal function via G-protein-coupled receptor 43 (GPR43)-mediated suppressing oxidative stress and NF-ƙB signaling pathway (Figure 5) [31, 111]. Administering L. casei Zhang orally can slow the decline in kidney function in patients with stage 3-5 CKD by modifying nicotinamide and SCFAs metabolism, thereby alleviating kidney damage and delaying the progression of renal decline [41]. Gemmiger formicilis is a chemoorganotrophic, gram-negative bacterium that synthesises butyrate via the acetyl coenzyme A pathway. The butyrate derivative sodium butyrate shows potential as a therapy for DKD, which suggests that Gemmiger may have nephroprotective properties [112]. Similar to humans, the diabetic rodent model exhibited dysbiosis, characterised by decreased α-diversity and increased abundance of Actinobacteria and Firmicutes, as well as a loss of SCFAs-producing bacteria such as Bacteroides, Rikenella, and Ruminococcus [113]. Perinatal propionate supplementation reverses CKD-induced hypertension in offspring of female rats. This protective effect correlates with increased plasma propionate and propionate-generating bacteria Clostridium spp. levels, enhanced expression of the renal GPR41 [114].
The molecular mechanism of microbial dysbiosis-mediated renal disease. During renal disease, pathogenic bacteria increase and beneficial bacteria decrease, causing metabolite disorders. These metabolites can activate the AHR receptor or NF-ƙB signalling pathway directly or indirectly, thereby exacerbating kidney disease. NPTX1, neuronal pentraxin 1; PKC, protein kinase C.
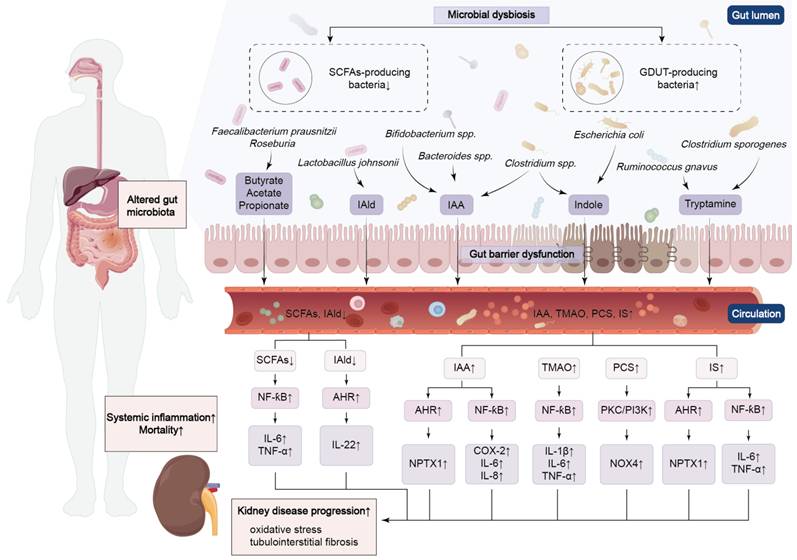
Reduced SCFAs are not merely correlates but causal drivers of renal fibrosis. In a preclinical mouse model of the transition from AKI to CKD, administration of butyrate and propionate both prevented disease progression and downregulated pro-fibrotic genes, demonstrating therapeutic potential independent of the intestinal microbiota [115]. In adenine-induced CKD mouse models, supplementation with butyrate effectively alleviated renal fibrosis, potentially by modulating nucleotide-binding oligomerization and domain-like receptor family protein 3 (NLRP3)-mediated pyroptosis through the Stimulator of interferon genes/NF-ƙB/p65 pathway [116]. Butyrate supplementation decreases the overexpression of TGF-β, which is induced by persistent, low-grade inflammation. It also suppresses the dilatation of glomerular and mesangial areas, oxidative damage and interstitial fibrosis in DKD mice through Nrf2-dependent mechanisms [31]. It is worth noting that the effects of SCFAs in inhibiting renal fibrosis and mesangial matrix accumulation are primarily mediated by GPR43 overexpression or GPR43 agonists. This confirms that SCFAs, particularly butyrate, may improve renal fibrosis through GPR43-mediated suppression of oxidative stress and NF-κB signaling [117]. Therefore, supplementing SCFAs may represent a promising direction for future research on renal fibrosis.
3.3.2. Tryptophan metabolites: AHR ligands
The accumulation of tryptophan metabolites in patients with renal failure is due to alterations in enzyme activity, gut microbial dysbiosis, and inefficient renal excretion, which ultimately resulting in an increase in tryptophan catabolites in intestine [67]. A study involving 63 patients with sepsis found that amino acid metabolism, which adjusts immunity and inflammation, was commonly dysregulated [118]. The biosynthesis of tryptophan and tyrosine was associated with the key molecular alterations within the multiomics network in sepsis. Phenylalanine metabolism was found to be associated with sepsis-related AKI [118]. It has been demonstrated that AHR activation in kidney disease is both tissue- and time-dependent. Using transgenic mouse models, Walker et al. showed that adenine-induced CKD not only leads to significant AHR activation in renal tubules, but also in hepatocytes, cardiac myocytes, and cardiac myocytes, hepatocytes, and microvasculature in the cerebral cortex. They also demonstrated that this activation pattern is closely correlated with serum IS levels [119]. In I/R-induced AKI, AHR activation in the renal tubules persists even after serum uremic toxin levels have decreased, suggesting the presence of other local injury-related mechanisms [119]. Intestinal bacterial translocation and low levels of endotoxemia, alongside decreased levels of proline, tryptophan, and tyrosine, have been observed in mice with renal I/R injury [96]. Bifidobacterium longum modulated macrophage inflammasome formation and mitochondrial fatty acid oxidation via histone deacetylase 3-mediated indole-3-carboxaldehyde, thereby counteracting intestinal I/R in mice [120]. Furthermore, Balestrieri et al. revealed that AHR activation in muscle exacerbates ischaemic pathology in CKD and that AHR activation in muscle is a key regulatory factor in ischaemic limb pathology in CKD [121]. Muscle-specific AHR knockout improved limb perfusion and preserved muscle mass and mitochondrial function, whereas constitutive AHR activation resulted in myopathy [121]. Therefore, gut-derived tryptophan metabolites that act as AHR ligands not only drive renal fibrosis, but also extra-renal complications of CKD.
A study of patients with stage 1-5 CKD found Lactobacillus johnsonii (L. johnsonii) supplementation attenuated renal lesions by restraining AHR signaling via augmented serum IAld levels [36]. A recent study revealed that the combination of AHR and IAld enhances the production of IL-22 by lamina propria lymphocytes, triggering the phosphorylation of transduction factor 3 and signal transduction factor. This stimulated the proliferation of mouse intestinal epithelial cells and repairs intestinal mucosal injury (Figure 5) [122]. Previous several studies indicated that IAld acted as a ligand of AHR and reduced levels of IL-1β, TNF-α, monocyte chemoattractant protein-1 (MCP-1), and cytochrome P450 family 1 subfamily A member 1 in mice [123, 124]. In adenine-induced CKD rats, serum IAld levels demonstrated a strong negative relationship with creatinine levels, confirming that L. johnsonii supplementation ameliorates renal lesions [36].
Gut microbiota that produces uremic toxin precursors were found to be more prevalent in CKD patients than in healthy controls [99]. Therefore, uremic toxins, including IS and IAA, accumulated progressively in the course of CKD [30, 125]. IS-mediated AHR binding to NF-ƙB p65 subunit, resulting in mutual inhibition of NF-ƙB and AHR, which inhibited the secretion of IL-6 and TNF-α in macrophages of CKD patients [126]. Serum IAA has been identified as a potential independent predictor of CVD and mortality in CKD patients, due to its ability to induce endothelial inflammation and oxidative stress, increase cyclooxygenase-2, IL-6, and IL-8 levels, and activate the inflammatory AHR/p38-mitogen-activated protein kinase/NF-ƙB pathway (Figure 5) [127]. Two species, Fusobacterium nucleatum and Eggerthella lenta, increased uremic toxins production in the adenine-induced CKD rats, with the former producing phenol and indole and the latter producing hippuric acid, which promote kidney damage [40]. Moreover, IAA and IS upregulate expression of neuronal pentraxin 1, which belongs to the inflammatory protein family, in an AHR-dependent manner (Figure 5) [128].
Recent studies have established causal links between tryptophan metabolites and renal fibrosis. Xie et al. showed that IS activated AHR to promote renal senescence and fibrosis via receptor gamma coactivator 1-α degradation, and AHR knockout abolished this effect [129].Intrarenal 1-methoxypyrene, an endogenous AHR agonist, accumulated progressively in obstructed kidneys and directly mediated tubulointerstitial fibrosis via the AHR. AHR knockdown or the use of specific AHR antagonists could attenuate 1-methoxypyrene-induced fibrosis [17]. Convergently, targeting Lactobacillus johnsonii in CKD patients and animal models increased serum IAld, a protective tryptophan metabolite that suppressed AHR signaling and reversed established kidney lesions [36]. A reduced relative abundance of Bifidobacterium animalis, Lactobacillus murinus, Lactobacillus johnsonii, Lactobacillus reuteri, and Lactobacillus vaginalis was found to be positively correlated with decreased tryptamine, indole-3-pyruvic acid, and IAld levels, and negatively correlated with increased IAA and indole-3-lactic acid levels in the serum of MN rats [39]. In summary, these findings suggest a causal relationship between tryptophan metabolites produced by the gut microbiota and the progression of renal fibrosis.
3.3.3. TMAO
TMAO was identified as a key metabolite linked to the AKI-to-CKD transition in both AKI patients and AKI-to-CKD transition mice [130, 131]. In vivo and in vitro studies confirmed that NOX2 activation was a critical regulator of TMAO-related transition from AKI to CKD [131]. TMAO has been shown to cause kidney damage and tubulointerstitial fibrosis, and elevated plasma concentrations of TMAO in CKD patients predict poor long-term survival [132, 133]. Among community-dwelling US adults, higher serial plasma TMAO levels were linked to a higher risk of incident CKD and a greater annual decline in kidney function [132]. The advent and progression of TMAO-mediated CKD principally comprise inflammatory mechanisms. A clinical study confirmed that DKD patients with higher plasma TMAO levels had poorer renal outcomes and a higher mortality rate. TMAO activated the NF-ƙB pathway in DKD patients, exacerbating systemic microinflammation and accelerating DKD progression [100]. TMAO increased renal scarring via TGF-β1 signaling pathways, and circulating TMAO levels were substantially reduced in patients receiving hypoglycemic drugs with reno-protective properties, such as GLP-1 receptor agonists [134]. Nondialysis patients with stage 3-5 CKD who exhibited rapid estimated glomerular filtration rate (eGFR) decline possessed a unique intestinal microbial composition, characterized by elevated α-diversity and an abundance of TMA-producing bacteria, such as Collinsella tanakaei and Desulfovibrio [135]. In vivo studies suggest that TMAO activates the NOD-like receptor protein 3 inflammasome, leading to the secretion of IL-18 and IL-1, and exacerbating renal inflammation and fibrosis (Figure 5) [31]. Targeting macrophage phenotype through gut-kidney axis improved renal fibrosis in mice. In both in vitro and in vivo culture experiments, TMAO stimulation was found to increased mRNA expression for IL-1β, IL-6, nitric oxide synthase, and TNF-α [136]. TMAO worsens kidney damage caused by hyperoxaluria in mice by activating the protein kinase R-like endoplasmic reticulum kinase/ROS pathway. This boosts apoptosis, autophagy, and inflammation, while also promoting calcium oxalate crystals accumulation in renal tubular cells [137]. These findings suggest that TMAO shows great potential as a therapeutic target for treating renal fibrosis, and we plan to develop strategies for treating renal disease by modulating TMAO levels.
3.3.4. PCS
Lactobacillus salivarius BP121 prevented cisplatin-induced AKI in rats by modulating the intestinal environment and reducing inflammation and oxidative stress, thereby inhibiting PCS and IS production, while regulating AMP-activated protein kinase- and Toll-like receptor 4 (TLR4)-dependent tight junction protein assembly [138]. With the advancement of CKD, phenylalanine, tyrosine, and tryptophan metabolism is enhanced within uremic toxin-related metabolic pathways, in contrast, proline and arginine metabolism is weakened [106]. PCS is a protein-binding uremic toxin that accelerates the deterioration of renal function in CKD patients and raises the risk of CVD. Microbiota-driven therapies, such as synbiotics or prebiotics, can reduce circulating PCS concentrations in patients with CKD [139]. A study of urine and serum samples taken from 90 DKD patients in different stages revealed that urinary PCS and serum metabolites such as butenoylcarnitine, arginine, and IS may serve as biomarkers of early DKD [140]. Freise et al. found that the effects of gut-derived uremic toxins (PCS, TMAO, and IS) on glycosaminoglycans involve the activation of the transcription factor NF-ƙB and of the phosphatidylinositol-3 kinase (PI3K)/protein kinase B pathway [141]. PCS activates ROS production and nicotinamide adenine dinucleotide phosphate oxidase through the protein kinase C and PI3K signalling pathways, triggering the expression of inflammatory cytokines in 5/6-nephrectomized rats (Figure 5) [142]. Moreover, Wu et al. also found that reduced Prevotella sp. 885 correlated with urea excretion, while elevated PCS inversely correlated with GFR [143].
PCS can directly induce renal tubular damage and fibrosis. Administration of PCS for four weeks in 5/6-nephrectomized rats caused significant tubular injury and oxidative stress [142]. Knockdown of NOX4 suppressed PCS-induced pro-oxidant effects, and probenecid blocked its toxicity, confirming that intracellular accumulation and NADPH oxidase activation are required for its profibrotic action [142]. Taken together, modulating gut-derived uremic toxins PCS may provide an additional effective therapeutic strategy for treating kidney disease.
3.3.5. Polyamines
Sepsis-induced AKI in mice was prevented by spermidine, which decreased NLRP3 inflammasome activation and IL-1β production [144]. In CKD rats, increased acrolein, spermidine, spermine, N1,N12-diacetylspermine, and N1-acetylspermidine were associated with an increase in Escherichia_Shigella, Clostridium_sensu_stricto, Parasutterella, and Enterorhabdus, while increased putrescine was linked to Bacteroides [145]. Mice with CKD following 5/6 nephrectomy exhibited reduced fecal output, with this constipation associated with suppressed ileal contractile responses. Spermine was found to suppress ileal contractile responses [146]. Taken together, microbiome-derived metabolites are altered in kidney disease and can influence the progression of the disease.
The development of renal fibrosis is influenced by polyamine metabolism. Causal evidence establishes polyamine metabolism as a determinant of renal fibrosis. Zahedi et al. demonstrated that ablation of polyamine catabolic enzymes significantly reduced cisplatin-induced renal injury and fibrosis, proving that enhanced polyamine catabolism drives fibrotic progression [147]. In contrast, Nakano et al. showed that spermidine activates Nrf2 and suppresses TGF-β1 signaling and oxidative stress, and spermidine treatment prevents fibrotic progression in arginase 2 knockout mice, establishing a protective causal pathway [148]. The expression of SMOX, a vital enzyme that regulates polyamine metabolism, is positively associated with renal fibrosis and function decline in CKD patients [149]. Luo et al. discovered that the SMOX/spermine axis could be a new, promising therapeutic approach for counteracting renal fibrosis, potentially by suppressing senescence and coordinating autophagy [149]. Therefore, polyamines influence the progression of renal fibrosis, and targeting gut microbiota may be an effective strategy for patients to slow the progression of renal disease. Many therapeutic strategies currently target this critical aspect.
3.3.5. Differential gut microbial and metabolite in renal disease
Studies have demonstrated significant differences in the dysbiosis of gut microbiota and metabolites between AKI and CKD, as well as between early and advanced stages of CKD. Plasma TMAO has been identified as significantly associated with an increased risk of AKI-to-CKD transition. NOX2 activation has been confirmed as a key regulator mediating TMAO-related AKI-to-CKD progression, both in vitro and in vivo [131]. Furthermore, plasma TMAO levels in CKD patients can predict long-term mortality and cardiovascular events, highlighting its prognostic value [150]. A large population-based cohort study demonstrated that PCS, IS, and phenylacetylglutamine were independently and inversely associated with eGFR, serving as early markers of renal function decline [151]. Propionate was substantially lower in advanced CKD and exhibited high discriminative power between patients with advanced CKD and non-CKD controls [143]. These investigations indicate that specific microbial by-products might act as early alerts for the advancement of AKI to CKD or as indicators for the severity of CKD.
In CKD, progressive alterations in gut microbial composition and metabolite profiles correlate with disease severity. Wu et al. found that the levels of six circulating metabolites and thirteen microbial species changed significantly from early to advanced CKD. Among them, Bacteroides eggerthii and Prevotella sp. 885 were the most effective features for classifying mild and advanced CKD, respectively, and Bacteroides eggerthii was particularly valuable for early diagnosis [143]. A core CKD-associated microbiota comprising seven genera and two species (Bacteroides eggerthii and Collinsella stercoris) was discovered to be highly correlated with CKD staging. Notably, Collinsella stercoris, Paraprevotella, and Pseudobutyrivibrio could distinguish early CKD from controls with better discriminatory performance than the conventional urine protein-to-creatinine ratio [107]. In the PREDIMED-Plus trial, lower abundances of the butyrate-producing genera Lachnospira and Lachnoclostridium were associated with CKD progression [152]. Recent studies revealed that patients with severe CKD harbored a higher enrichment of uremic toxin-producing species compared with those with moderate CKD. Furthermore, toxin-producing bacterial species gradually increased over time, whereas microbial diversity progressively decreased [153].
Collectively, these findings indicate that gut microbial dysbiosis evolves dynamically with the progression of CKD, characterized by the enrichment of uremic toxin-producing bacteria and the depletion of SCFAs-producing bacteria, accompanied by differential changes in metabolites such as TMAO. Thus, these factors hold potential as biomarkers for monitoring disease progression.
4. The therapeutic strategies in renal disease by modulating gut microbial-derived metabolites
The understanding of underlying molecular mechanisms in disease aims to discover novel therapeutic agents and strategies. The mentioned-above studies indicate that the changes in gut microbial-derived metabolites in kidney disease including both AKI and CKD. Based on the elucidation of underlying molecular mechanisms, increasing studies suggest that a number of therapeutic strategies, such as dialysis (hemodialysis, peritoneal dialysis and colonic dialysis), KT, natural product interventions, microecological therapies (probiotics, prebiotics and synbiotics), and dietary interventions, enhanced kidney function and improved renal injury by regulating gut microbial-derived metabolites [154, 155]. Therefore, targeting gut microbiota has been demonstrated as an effective and promising therapy for treating kidney diseases.
4.1 Regulating gut microbial-derived metabolites by renal replacement therapies
In the last several decades, extensive clinical data have shown renal replacement therapies including KT and dialysis, such as hemodialysis, peritoneal dialysis and colonic dialysis, is the final treatment for patients with ESRD and uremia that improve life quality [156-158]. Increasing publications have revealed that these therapies can affect the levels of the gut microbial-derived metabolites, such as IS, PCS and TMAO and regulated pro-inflammatory cytokines and AHR signalling pathway [91, 159].
4.1.1 Regulating microbial-derived metabolites by hemodialysis and peritoneal dialysis
Hemodialysis (HD) is a commonly used dialysis method for patients with ESRD and uremia worldwide [160-162], while peritoneal dialysis (PD) is also widely used for the long-term treatment of ESRD, accounting for 9% of all renal replacement therapy and 11% of all dialysis globally [154, 163]. PD is a renal replacement therapy for ESRD, and is now commonly used to achieve target clearances for creatinine or urea [164]. Lin et al. demonstrated that distinct gut microbial composition was linked to an elevated mortality risk in HD patients, and that the abundance of Anaerostipes and Succinivibrio, two SCFAs-producing bacteria, was markedly lower in non-survivors [155]. Changes in the microbiome of CKD patients are phase-dependent, being most pronounced in HD, with a decrease in saccharolytic bacteria, such as Bifidobacterium, and an increase in proteolytic bacteria, such as Citrobacter [91]. HD patients showed lower plasma tryptophan levels but higher IS and five kynurenine metabolites, including anthranilic acid, kynurenic acid, kynurenine, xanthurenic acid, and 3-hydroxykynurenine [91]. Since kynurenine and IS can activate AHR, serum-induced AHR activity was higher in HD patients than healthy controls [91]. HD patients have higher levels of serum IS and PCS than healthy controls. These protein-bound uremic toxins are difficult to remove by dialysis and are key factors in the development of CVD [165]. TMAO, a gut microbial metabolite, raises the risk of peritoneal inflammation and peritonitis in PD patients. In vitro study demonstrated that TMAO directly triggered necrosis of primary peritoneal mesothelial cells, alongside elevated production of pro-inflammatory cytokines, including IL-6, IL-1β, TNF-α, and MCP-1. Furthermore, TNF-α-induced P-selectin generation in mesothelial cells was considerably increased by TMAO [159]. Both HD and PD patients exhibited increased levels of proinflammatory cytokines, such as TNF-α and IL-6 [155, 166]. IL-6 has been revealed to affect the structural and inflammatory properties of the peritoneal membrane [166]. HD patients exhibit myeloid cells alters, shown by a shift from classical to non-classic and proinflammatory intermediate monocytes, which efficiently produce TNF-α [91]. Collectively, these findings suggest that both hemodialysis and peritoneal dialysis affect the gut microbial-derived metabolites in CKD patients.
4.1.2 Regulating microbial-derived metabolites by colonic dialysis
Colonic dialysis has long been employed in China to remove intestinal-derived uremic toxins and delay CKD progression. The injection of an osmotically balanced solution into the colon during colonic dialysis flushes out persistent stool and intestinal-derived uremic toxins [167]. Colonic dialysis ameliorated renal function in patients with stage 3-5 CKD and increasesd the abundance of anaerobic bacteria producing SCFAs in CKD patients, including Bifidobacterium, Bacteroides, and Collinsella, while reducing the abundance of microbes that possess urease and generate PCS or IS, including unclassified Enterobacteriaceae [167, 168]. A 20-year-old female patient with ESRD underwent colonic dialysis via a Malone antegrade continent enema stoma for two years. Throughout this period, her serum blood urea nitrogen (BUN) and creatinine levels remained consistently low, suggesting that this approach could be a promising alternative to renal replacement therapy [169]. Considerable decreases in creatinine and BUN levels were observed following colonic dialysis in uremic rats that had undergone left partial nephrectomy and right nephrectomy [170]. Clinical evidence suggests that a combination of colonic dialysis and traditional Chinese medicine enemas can treat chronic renal failure by reducing Scr levels [171]. Colonic dialysis using Chinese medicine altered creatinine decomposition by gut bacteria in uremic rats, reducing Lactobacillus and Bifidobacterium abundance and increasing E.coli abundance in the colon [172]. Colonic dialysis with Gubenxiezhuo significantly improved apoptosis and inflammatory infiltration in uremic rats, reduced hemorrhage, and attenuated the expression levels of C-reactive protein, IL-6, and IL-1β [173]. Therefore, colonic dialysis may be a promising approach to treating kidney disease in the future.
4.1.3 Regulating microbial-derived metabolites by KT
Compared to dialysis, KT offers a higher survival rate and improves life quality, making it an advantageous treatment option for patients with ESRD [174-176]. Compared to PD or HD, KT reduced levels of several uremic toxins and also reduced the risk of CVD [67, 177]. Swarte et al. found that a lower abundance of four butyrate-producing bacteria, including Eubacterium hallii, Firmicutes bacterium CAG 83, Faecalibacterium prausnitzii, and Gemmiger formicilis, was linked to a higher risk of death. KT patients had lower butyrate-producing bacteria abundance than healthy controls. Lower levels of these bacteria also linked to poorer health-related quality of life in KT patients [178]. Metabolites like kynurenine, polyunsaturated fatty acid, uric acid, creatinine were identified as differential serum metabolites in acute post-transplant graft rejection [179]. Although TMAO levels in KT recipients declined sharply, they did not reach the levels seen in healthy controls. Furthermore, an increased mortality risk was independently associated with elevated plasma TMAO concentrations in KT patients [180]. A high-fiber diet or sodium acetate supplementation altered the microbiome, modifying alloimmunity in a mouse KT model. This generated tolerance dependent on GPR43 and Tregs [181]. Collectively, these findings emphasize complex interplay between metabolic dysregulation, gut microbiota, and overall health in patients receiving renal replacement therapy.
4.2 Regulating gut microbial-derived metabolites in renal disease by natural products
Traditional Chinese medicines have been long practiced and are commonly recognised as an essential treatment for various kidney diseases including AKI and CKD [182-189]. Recent studies emphasize natural products improved AKI and CKD by regulating the dysbiosis of gut microbiota and the disorder of gut microbial-derived metabolites [36, 39, 190].
4.2.1 Regulating gut microbial-derived metabolites in AKI by natural products
A larger body of research study has demonstrated that natural products exhibit a significant therapeutic effect in ameliorating AKI [191-195]. Studies have demonstrated that natural compounds exhibit obvious therapeutic effects on animal models with renal disease. Total alkaloids of Aconitum carmichaelii Debx (ACA) retarded cisplatin-induced AKI by decreasing relative abundance of Escherichia-Shigella, Ruminococcus, and Clostridium, while increasing Bacteroides, Ligilactobacillus, Desulfovibrio, and Anaerotruncus [196]. This were accompanied by reducing uremic toxins, and elevating serum levels of SCFAs that were related to the glutathione and tryptophan metabolism, decreasing serum levels of inflammatory factors, such as IL-1β, IL-6, and TNF-α, while restoring levels of superoxide dismutase, catalase and glutathione, and decreasing monoester diterpene alkaloid levels in the kidney by inhibiting NF-κB pathway and increasing Nrf2/HO-1 pathway [196]. Treatment with loganetin ameliorated rhabdomyolysis-induced AKI by inhibiting TLR4 activity and suppressing NF-ƙB and c-Jun N-terminal kinase/p38 pathways [197]. Natural polysaccharides, important bioactive components derived from traditional Chinese medicine, are recognized for their therapeutic potential in treating various diseases by improving gut microbial dysbiosis [198-200]. The latest study revealed that alginate oligosaccharide inhibited renal I/R injury by upregulating mannose receptor type C1 expression and suppressing TLR4/myeloid differentiation primary response 88/NF-ƙB/IL-1β inflammatory pathway [194]. Moreover, selenium-enriched crude polysaccharide (SeRRP) ameliorated cadmium-induced AKI by elevating relative abundance of Muribaculaceae, Ruminococcaceae, and Lachnospiraceae [201]. In addition, treatment with 5,2'-dibromo-2,4',5'-trihydroxydiphenylmethanone alleviated acute pyelonephritis by inhibiting inflammatory mediators and regulating immune responses [202] (Table 1). Therefore, these findings demonstrate the potential for natural products to affect gut-kidney axis, thereby mitigating AKI positively.
Treatment of natural products in patients and animal models with AKI.
| Drugs | Disease | Dose | Microbial dysbiosis | Molecular mechanism | Refs |
|---|---|---|---|---|---|
| ACA | Cisplatin-induced AKI mice | 5, 10 mg/kg | Increased Bacteroides and Ligilactobacillus abundance while decreasing Clostridium abundance. | Restored glutathione and tryptophan metabolism and improved IƙB/NF-ƙB and Keap1/Nrf2 pathways. | [196] |
| Loganetin | AKI mice | 2, 18 mg/kg | - | Inhibited TLR4 activity and suppressed NF-ƙB and JNK/p38 pathways. | [197] |
| Alginate oligosaccharide | I/R-induced AKI rats | 100 mg/kg | - | Inhibited TLR4/MyD88/NF-ƙB/IL-1β pathway. | [194] |
| SeRRP | Cadmium-induced AKI mice | 100 mg/kg | Increased Muribaculaceae, Ruminococcaceae and Lachnospiraceae abundance. | Modulated gut microbiota and antioxidant activity. | [201] |
Abbreviations: JNK, c-Jun N-terminal kinase; Keap1, kelch-like ECH-associated protein 1; MyD88, myeloid differentiation primary response 88.
4.2.2 Regulating gut microbial-derived metabolites in CKD by natural products
Compared with AKI, numerous research has reported that natural products improved CKD and inhibited renal fibrosis [203-209]. Increasing evidence has shown that traditional Chinese medicines ameliorated patients with CKD by reshaping microbial dysbiosis [210-212]. Accumulated studies have demonstrated that natural products can ameliorate renal injury in CKD patients [213-215]. For instance, a randomized controlled trial demonstrated that Yi-Shen-Hua-Shi enriched SCFA-producing bacteria and reduced uremic toxin-producing bacteria in CKD patients, resulting in decreased proteinuria and improved renal function [210]. Zicuiyin decoction increased relative abundance of Prevotellaceae and Lactobacillaceae and decreased Clostridiaceae, Enterobacteriales, and Micrococcaceae in DKD patients [216]. Jin Gui Ren Qi Pill and western medicine enhanced the abundance of Prevotella_7 within the gut microbiota of DKD patients, exhibiting superior efficacy in alleviating clinical symptoms compared to western medicine treatment alone [217]. Modified Zexie decoction could enhance microbial diversity, increase SCFAs, alleviate inflammation, improve renal function, and alleviate phlegm-dampness hypertension in patients by regulating the gut-immune-kidney axis [218]. It was ascertained that Shen-Shuai-Ning granule could reduce the total serum IS concentration in PD patients at weeks 4, 8, and 12. Nevertheless, further large-scale, long-term clinical trials were warranted for validation [219]. Fushen granule showed beneficial therapeutic effects in PD patients by increasing relative abundance of Rothia, Megamonas, and Bacteroides, which is closely associated with improved amino acid metabolism (Table 2) [212].
Treatment of natural products including traditional Chinese medicine in patients and animal models with CKD.
| Drugs | Disease | Dose | Microbial dysbiosis | Metabolites or effects | Mechanism | Refs |
|---|---|---|---|---|---|---|
| Yi-Shen-Hua-Shi | CKD patients | 1 bag/ 3 times/day | Increased Lachnoclostridium, Lachnospiraceae and Sutterella abundance while decreased Clostridium innocuum and Eggerthella abundance. | Reduced 24-h proteinuria level. | Modulated gut microbiota and regulated vitamin, lipid and glycan metabolism. | [210] |
| Zicuiyin Decoction | DKD patients | 150 mL/day | Increased Lactobacillaceae abundance while decreased Clostridiaceae abundance. | Decreased serum creatinine level. | - | [211] |
| Jin Gui Ren Qi Pill | DKD patients | 5 g/ 2 times/day | Increased Prevotella_7 abundance. | Reduced IL-2 level. | Modulated gut microbiota. | [217] |
| Modified Zexie decoction | Hypertension patients | 150 mL/2 times/day | Increased Streptococcus and Prevotella abundance while decreased Escherichia and Agathobaculum abundance. | Increased acetate, propionate and butyrate levels. | Regulated gut-immune-kidney axis and enhancing microbial diversity. | [218] |
| Shen-Shuai-Ning granule | PD patients | 5 g/3 times/day | - | Reduced IS level. | Decreased uremic toxins. | [219] |
| Fushen Granule | PD patients | 18 g/2 times/day | Increased Rothia, Megamonas and Bacteroides abundance. | Reduced serum creatinine and urea levels. | Improving carbohydrate and amino acid metabolism. | [212] |
| Fisetin | Hyperuricemia-induced CKD mice | 50,100 mg/kg | - | Decreased tryptophan metabolites. | Modulated tryptophan metabolism and AHR activation. | [220] |
| Berberine | Adenine-induced CKD rats | 100 mg/kg | Increased butyric acid-producing bacteria abundance. | Increased butyric acid level while reduced TMAO level. | Inhibited tyrosine metabolism. | [221] |
| Punicalagin | High-fat diet-induced DKD mice | 50, 100 mg/kg | Increased Lachnospiraceae abundance. | Increased SCFA level while reduced LPS and DAO levels. | Improving diabetic renal injury through gut-kidney axis. | [222] |
| Lobetyolin, luteolin, and rutin | Streptozotocin-induced DKD rats | 75 mg/kg | Increased Ruminococcus and Barnesiella abundance while decreased Oscillospira abundance. | Increased SCFA level while reduced LPS level. | Modulated gut microbiota. | [223] |
| ABPW1 | Streptozotocin-induced DKD mice | 300 mg/kg | Increased Bacteroide abundance while decreased Alistipes, Faecalibaculum, Rikenella, and Laedolimicola abundance. | Increased SCFA level. | Modulated gut microbiota. | [224] |
| OFI-F | Streptozotocin-induced DKD mice | 300, 500 mg/kg | Increased Akkermansia and Bacteroides abundance while decreased Alistipes, Helicobacter, and Desulfovibrio abundance. | Increased SCFA level. | Regulated gut microbiota and promoting SCFAs production. | [225] |
| Polygonum cuspidatum | Hyperuricemia patients, potassium oxonate and adenine-induced rats | 2, 20, 200 mg/kg | Increased Lactobacillus and Candidatus_Arthromitus abundance while decreased Pseudomonas, Helicobacter, and Romboutsia abundance. | Decreased leucine, phenylalanine, and tyrosine levels. | Rectifying amino acid metabolism disorders by modulated gut microbiota dysbiosis. | [226] |
| Jinlida | DKD mice | 1.75, 3.5 mg/kg | Increased Paramuribaculum intestinale abundance. | Increased pyridoxamine and pyridoxine levels. | Reshaping gut microbiota and suppressed AGEs-RAGE axis. | [227] |
| Guizhi Fuling pills | Carbon tetrachloride-induced mice | 125 mg/kg | Increased Bacteroidales-S24-7-group abundance while decreased Lactobacillus abundance. | Decreased IL-6, IL-1β, and TNF-6 levels. | Regulated NLRP3 inflammasome, Nrf2 pathway, and gut microbial composition. | [228] |
| Simiao decoction | Uricase-knockout rats | 3, 6 g/L drugs in drinking water | Increased Lactobacillus_intestinalis abundance. | Decreased IL-6, IL-1β and MCP-1 levels and downregulated uremic toxins including IS, PCS, and methylguanidine. | Modulated gut dysbiosis and decreased uremic toxins. | [229] |
| Fucoidan | UUO-induced mice | 25, 50, 100 mg/kg | Increased Akkermansia muciniphila abundance. | Increased SCFA level. | Inhibited NEU1-TLR4-NFƙB-mediated inflammation. | [232] |
| DSP | Hyperuricemia mice | 100, 200, 300 mg/kg | Increased Akkermansia and Lactobacillus abundance while decreased Bacteroides abundance. | Reduced TNF-α, IL-1β, IL-6, and LPS levels. | Inhibited NF-ƙB pathway by gut-kidney axis. | [233] |
| RG-I | Glyoxylate-induced rats | 10, 20, 40 mg/kg | Increased Enterobacterales, Lactobacillus, and Mediterraneibacter abundance. | Increased taurine level while reduced TNF-α level. | Regulated gut microbiota and suppressed TNF-α expression in kidneys. | [234] |
Abbreviations: ABPW1, a graminan type fructan from Achyranthes bidentata; APSP, extract mixture of Andrographis paniculata (Burm. f.) Wall. ex Nees and Syzygium polyanthum (Wight.) Walp leaves; BSA, barleriside A; DAO, diamine oxidase; DSP, Dioscorea septemloba polysaccharides; HMF, 5,6,7,8,3',4'-hexamethoxyflavone; NEU1, neuraminidase-1; OFI-F, flavonoids derived from Opuntia ficus-indica fruit; RG-I, rhamnogalacturonan I from Polygonum aviculare L.; UUO, unilateral ureteral obstruction; 16(R)-HETE, 16(R)-hydroxy-5,8,11,14-eicosatetraenoic acid.
Accumulating evidence has demonstrated that natural products play a crucial role in preserving intestinal barrier integrity and regulating microbial dysbiosis in CKD. For instance, fisetin protected against hyperuricemia-induced CKD by modulating gut microbiota-derived tryptophan metabolites and activating AHR signalling pathway [220]. Moreover, berberine inhibited tyrosine metabolism within gut microbiota dysbiosis and reduced nephrotoxic TMAO levels [221]. Previous studies have revealed that a number of natural components mitigated DKD by increasing SCFA levels. For example, punicalagin increased SCFA concentrations and inhibited serum diamine oxidase and LPS levels, possibly alleviating renal injury through gut-kidney axis [222]. Rutin, lobetyolin and luteolin, the primary active components of the Hong Guo Ginseng Guo, mitigated DKD by regulating gut microbiota and inhibiting NLRP3 inflammasome [223]. A graminan type fructan from Achyranthes bidentata alleviated renal injury in DKD mice by restoring barrier function, modulating microbial dysbiosis, and increasing SCFAs levels [224]. In addition, flavonoids derived from Opuntia ficus-indica fruit attenuated renal injury in DKD mice by altering gut microbiota, suppressing LPS-induced inflammation, and promoting SCFA production [225]. In hyperuricemic rats, Polygonum cuspidatum rectified amino acid metabolic disorders including those of tyrosine, leucine, and phenylalanine by modulating gut microbiota dysbiosis [226]. Jinlida reshaped the gut microbiota, elevated pyridoxamine and pyridoxine levels, inhibited the advanced glycation end-products (AGEs)-the receptor for AGEs (RAGE) axis, and thereby suppressed downstream Smad2-mediated fibrosis and NF-κB-driven inflammation, exerting a renoprotective effect [227]. Guizhi Fuling pills might ameliorate renal injury in mice by regulating the NLRP3 inflammasome, Nrf2 pathway, and gut microbial composition [228]. Simiao decoction alleviated hyperuricemia-induced renal injury by modulating gut microbial metabolism of tyrosine and tryptophan, thereby decreasing uremic toxins such as IS and PCS [229]. Polysaccharides derived from natural products have a broad spectrum of sources and possess various pharmacological effects [230, 231]. Fucoidan improved renal fibrosis by Akkermansia muciniphila-produced acetate, which subsequently inhibited neuraminidase-1/TLR4/NF-κB pathway [232]. In addition, Dioscorea septemloba polysaccharides alleviate renal disease and hyperuricemia by improving intestinal barrier function injury, gut microbial dysbiosis and lipid metabolism disorder (Table 2) [233]. Rhamnogalacturonan I from Polygonum aviculare L. regulated the gut microbiota and promoted taurine production, thereby inhibiting TNF-α expression, relieving local inflammation, and exerting anti-nephrolithiatic effect [234]. Taken together, natural products are a potentially therapeutic approach for treating kidney disease.
4.3 Regulating gut microbial-derived metabolites in CKD by microecological therapy
A growing body of evidence shows that microbial therapeutics including probiotics, prebiotics, and synbiotics [35, 154, 235] are the most commonly used method for regulating gut microbiota in diverse diseases [236, 237]. Supplementation with Faecalibacterium prausnitzii attenuated renal inflammation, renal insufficiency, and decreased serum levels of numerous uremic toxins in CKD mice, which was partially attributed to butyrate-mediated renal GPR-43 signaling [111]. Supplementing with probiotic Bifidobacterium animalis reduced relative abundance of Fusobacterium nucleatum and Eggerthella lenta, decreased uremic toxin levels and slowed the progression of kidney disease in CKD rats [40]. Supplementation with the probiotic DM02, comprising L. plantarum and L. reuteri, reduced IS and E. coli levels in patients with stage 3-5 CKD, attenuated heart failure progression in CKD rats, and lowered levels of nicotinic acid, IS, and N-acetylserotonin [238]. In 5/6 nephrectomy mice, Lactobacillus rhamnosus L34 alleviated uremia-induced systemic inflammation by decreasing uremic toxins such as IS and TMAO and gut leakage, thereby delaying CKD progression [239]. Additionally, supplementation with β-glucan prebiotic in CKD patients with stage 3 to 5 reduced colon-derived uremic toxins PCS, IS, and p-cresyl glucuronide levels, and showed a trend toward shifting the gut microbiota from a Bacteroides 2 to Prevotella enterotype [240]. Administering a prebiotic mixture of oligofructose and inulin alleviated renal function and frailty among the older population, while increasing the levels of alanine, histidine, and methionine. Within the microbiota, Dialister emerged as a key species, exhibiting positive correlations with indole-3-lactic acid, L-tryptophan, indole, and L-alanine [241]. Adenine-induced CKD rats supplemented with prebiotic gum acacia exhibited reduced Tenericutes, Verrucomicrobia, Proteobacteria, and Actinobacteria abundance, slowing down CKD progression by enhancing butyrate production [242]. The combination of Lactiplantibacillus plantarum MPB-65 and epicatechin as a synbiotic supplement mitigated hyperuricaemia by regulating the gut-kidney axis. It increased SCFAs levels, reduced serum BUN, uric acid, and creatinine levels, and inhibited hepatic xanthine oxidase activity [243].
Despite the promising benefits, the application of microecological therapies in CKD patients also carries potential risks and side effects that warrant careful consideration. Adverse events were reported infrequently and were mild in nature in patients with stage 1 to 5 CKD who were treated with synbiotics, prebiotics, or probiotics, and included abdominal pain, diarrhoea, flatulence, and nausea [244]. However, a meta-analysis of critically ill adult patients showed that probiotic and synbiotic administration was associated with an increased risk of adverse events, primarily bacteremia and intestinal ischemia [245].
In summary, although probiotics, prebiotics, and synbiotics show promise as additional therapies for kidney disease, their use should be determined following careful consideration of the patient's immune status, disease stage, and any other medications they are taking.
4.4 Regulating gut microbial-derived metabolites in CKD by dietary interventions
Compared to a low-fat diet rich in complex carbohydrates, long-term consumption of a Mediterranean diet enriched with extra-virgin olive oil protected renal function and slowed eGFR progression in DKD patients [246]. For instance, oat dietary fiber enhanced SCFAs levels in CKD mice, rectify intestinal dysbiosis, diminished uremic toxins, and alleviated renal fibrosis through NF-ƙB/TGF-β signaling pathway [247]. In summary, these strategies represent great promise as therapeutic management options for kidney disease.
5. Conclusion, perspective and challenges
Increasing evidence has indicated that there is a bidirectional relationship between kidney and gut microbiota. Maintaining homeostasis between the gut microbiota and the host is crucial for preventing renal fibrosis. The function and composition of gut microbiota are influenced by the uremic milieu. At the same time, an increase in microinflammation and uremic toxins derived from gut microbiota is linked to ecological disruption, both of which contribute to the progression of renal disease. Microbial-derived metabolites, including SCFAs and uremic toxins, modulate immune cells, control inflammatory mediators, and mediate the gut-mucosal barrier. Ultimately, they shape the overall environment, either disrupting or promoting the process of renal fibrosis. Using dialysis (hemodialysis, peritoneal dialysis and colonic dialysis), KT, natural product interventions, microecological therapies (probiotics, prebiotics and synbiotics), and dietary interventions to target the gut microbiota is a potential therapy for kidney disease, but it still faces several new challenges. Further research is required to clarify the pharmacological effects of significantly modified bacterial species and their metabolites in patients with kidney disease. Furthermore, it is vital to grasp the underlying molecular mechanisms of microbiological dysbiosis associated with kidney disease, which are triggered by microbial-derived metabolites. To deepen our comprehension of the physiological functions of gut microbiota and unravel the pathophysiology and etiology of kidney disease, sequencing, bioinformatics, and untargeted metabolomics technologies must be integrated. Further research is needed to investigate the underlying mechanisms of using natural products, probiotics, and prebiotics to restore homeostasis to the gut-kidney axis as a potential therapeutic strategy and to examine its clinical application. Although significant progress has been made in current research, the field still faces many challenges and opportunities. Notably, the deep integration of metagenomics with other emerging technological fields is providing unprecedented insights into the gut microbiota-kidney axis. Multi-omics integration strategies, which combine metagenomics, metatranscriptomics, metaproteomics, and metabolomics, enable the systematic analysis of interactions between the gut microbiota and the host at multiple levels, including composition, function, and metabolites. They also allow the systematic analysis of microbial functional and metabolite networks. Single-cell sequencing and spatial transcriptomics reveal heterogeneous responses of renal and intestinal cells to microbial metabolites. Furthermore, engineered strategies such as gene-edited probiotics, synthetic microbiota and phage therapy provide new opportunities for the precise regulation of harmful metabolites. However, the clinical translation of these technologies is hindered by challenges such as insufficient standardisation, high costs, poor model interpretability and difficulties in validating causal relationships. In the future, large-scale, multicentre, longitudinal follow-up cohorts should be established to integrate multi-omics and clinical data. This will facilitate progress from association to causation and from understanding mechanisms to translating them into practice. It will also enable precise, microbiome-targeted interventions.
Acknowledgements
Funding
This study was supported by National Natural Science Foundation of China (Nos. 82474062, 82274192 and 82274079), Zhejiang Provincial Natural Science Foundation (No. LHZSZ25H270001) and Shaanxi Key Science and Technology Plan Project (No. 2023-ZDLSF-26).
Competing Interests
The authors have declared that no competing interest exists.
References
1. Anandh U, Anders HJ, Bacchetta J, Johnson DW, Luyckx V, Remuzzi G. et al. Two decades of nephrology research: progress and future challenges. Nat Rev Nephrol. 2025;21:727-35
2. Blum MF, Neuen BL, Grams ME. Risk-directed management of chronic kidney disease. Nat Rev Nephrol. 2025;21:287-98
3. Huang Y, Ning X, Ahrari S, Cai Q, Rajora N, Saxena R. et al. Physiological principles underlying the kidney targeting of renal nanomedicines. Nat Rev Nephrol. 2024;20:354-70
4. Miguel V, Shaw IW, Kramann R. Metabolism at the crossroads of inflammation and fibrosis in chronic kidney disease. Nat Rev Nephrol. 2025;21:39-56
5. Abbad L, Esteve E, Chatziantoniou C. Advances and challenges in kidney fibrosis therapeutics. Nat Rev Nephrol. 2025;21:314-29
6. Peek JL, Wilson MH. Cell and gene therapy for kidney disease. Nat Rev Nephrol. 2023;19:451-62
7. Bensouna I, Doreille A, Dancer M, Lebre AS, Robert T, Mesnard L. Nephrogenomics, precision medicine and the role of genetic testing in adult kidney disease management. Nat Rev Nephrol. 2025;21:597-612
8. Huang R, Fu P, Ma L. Kidney fibrosis: from mechanisms to therapeutic medicines. Signal Transduct Target Ther. 2023;8:129
9. Zhang X, Zhang Y, Liu Y. Fibroblast activation and heterogeneity in fibrotic disease. Nat Rev Nephrol. 2025;21:613-32
10. Meng XM, Wang L, Nikolic-Paterson DJ, Lan HY. Innate immune cells in acute and chronic kidney disease. Nat Rev Nephrol. 2025;21:464-82
11. Zhao H, Guo J. Macrophages in focus: key drivers and therapeutic opportunities in diabetic kidney disease. Int J Biol Sci. 2025;21:4647-62
12. Wang YB, Li T, Wang FY, Yao X, Bai QX, Su HW. et al. The dual role of cellular senescence in macrophages: unveiling the hidden driver of age-related inflammation in kidney disease. Int J Biol Sci. 2025;21:632-57
13. Feng L, Feng YY, Ren Q, Fu P, Ma L. Mesangial cells in diabetic kidney disease: from mechanisms to therapeutic implications. Int J Biol Sci. 2025;21:4762-81
14. De Gregorio V, Barua M, Lennon R. Collagen formation, function and role in kidney disease. Nat Rev Nephrol. 2025;21:200-15
15. Miao H, Wang YN, Su W, Zou L, Zhuang SG, Yu XY. et al. Sirtuin 6 protects against podocyte injury by blocking the renin-angiotensin system by inhibiting the Wnt1/β-catenin pathway. Acta Pharmacol Sin. 2024;45:137-49
16. Liu T, Li Q, Jin Q, Yang L, Mao H, Qu P. et al. Targeting HMGB1: a potential therapeutic strategy for chronic kidney disease. Int J Biol Sci. 2023;19:5020-35
17. Cao G, Miao H, Wang YN, Chen DQ, Wu XQ, Chen L. et al. Intrarenal 1-methoxypyrene, an aryl hydrocarbon receptor agonist, mediates progressive tubulointerstitial fibrosis in mice. Acta Pharmacol Sin. 2022;43:2929-45
18. Kishi S, Nagasu H, Kidokoro K, Kashihara N. Oxidative stress and the role of redox signalling in chronic kidney disease. Nat Rev Nephrol. 2024;20:101-19
19. Hong Q, Kim H, Cai GY, Chen XM, He JC, Lee K. Modulation of TGF-β signaling new approaches toward kidney disease and fibrosis therapy. Int J Biol Sci. 2025;21:1649-65
20. Bloom PP, Garrett WS, Penniston KL, Winkler MH, Hazen SL, Agudelo J. et al. Microbiota and kidney disease: the road ahead. Nat Rev Nephrol. 2025;21:702-16
21. Miao H, Zhang YM, Yu XY, Zou L, Zhao YY. Membranous nephropathy: systems biology-based novel mechanism and traditional Chinese medicine therapy. Front Pharmacol. 2022;13:969930
22. Zhong Q, Zeng J, Li Y, Zhang H, Lin T, Song T. Senescent renal tubular cells derived extracellular vesicles transported miR-20a and miR-21 induced macrophage-to-myofibroblast transition in renal fibrosis after ischemia reperfusion injury. Int J Biol Sci. 2025;21:940-54
23. Liu J, Xiong Y, Mo H, Niu H, Miao J, Shen W. et al. MicroRNA-29b plays a vital role in podocyte injury and glomerular diseases through inducing mitochondrial dysfunction. Int J Biol Sci. 2024;20:4654-73
24. Ji JL, Shi HM, Li ZL, Jin R, Qu GT, Zheng H. et al. Satellite cell-derived exosome-mediated delivery of microRNA-23a/27a/26a cluster ameliorates the renal tubulointerstitial fibrosis in mouse diabetic nephropathy. Acta Pharmacol Sin. 2023;44:2455-68
25. Wang YN, Zhang ZH, Liu HJ, Guo ZY, Zou L, Zhang YM. et al. Integrative phosphatidylcholine metabolism through phospholipase A(2) in rats with chronic kidney disease. Acta Pharmacol Sin. 2023;44:393-405
26. Sharma K, Hansen J, Susztak K, Eberlin L, Anderton CR, Alexandrov T. et al. Spatial metabolomics and multiomics integration for breakthroughs in precision medicine for kidney disease. Nat Rev Nephrol. 2025;22:152-64
27. Wang Y, Liu T, Wu Y, Wang L, Ding S, Hou B. et al. Lipid homeostasis in diabetic kidney disease. Int J Biol Sci. 2024;20:3710-24
28. Liu HJ, Miao H, Yang JZ, Liu F, Cao G, Zhao YY. Deciphering the role of lipoproteins and lipid metabolic alterations in ageing and ageing-associated renal fibrosis. Ageing Res Rev. 2023;85:101861
29. Li XJ, Fang C, Zhao RH, Zou L, Miao H, Zhao YY. Bile acid metabolism in health and ageing-related diseases. Biochem Pharmacol. 2024;225:116313
30. Liu LM, He XF, Zhang YL, Zhou JT, Miao H, Zhao YY. Gut microbiota and renal fibrosis: novel mechanistic insights and therapeutic potential. Acta Pharmacol Sin. 2026
31. Li XJ, Shan QY, Wu X, Miao H, Zhao YY. Gut microbiota regulates oxidative stress and inflammation: a double-edged sword in renal fibrosis. Cell Mol Life Sci. 2024;81:480
32. Miao H, Zhang SJ, Wu X, Li P, Zhao YY. Tryptophan metabolism as a target in gut microbiota, ageing and kidney disease. Int J Biol Sci. 2025;21:4374-87
33. Si ZL, Wang HY, Wang T, Cao YZ, Li QZ, Liu K. et al. Gut Bacteroides ovatus ameliorates renal fibrosis by promoting the production of HDCA through upregulation of Clostridium scindens. Cell Rep. 2024;43:114830
34. Zhou W, Wu WH, Si ZL, Liu HL, Wang H, Jiang H. et al. The gut microbe Bacteroides fragilis ameliorates renal fibrosis in mice. Nat Commun. 2022;13:6081
35. Krukowski H, Valkenburg S, Madella AM, Garssen J, van Bergenhenegouwen J, Overbeek SA. et al. Gut microbiome studies in CKD: opportunities, pitfalls and therapeutic potential. Nat Rev Nephrol. 2023;19:87-101
36. Miao H, Liu F, Wang YN, Yu XY, Zhuang S, Guo Y. et al. Targeting Lactobacillus johnsonii to reverse chronic kidney disease. Signal Transduct Target Ther. 2024;9:195
37. Wang YN, Miao H, Yu XY, Guo Y, Su W, Liu F. et al. Oxidative stress and inflammation are mediated via aryl hydrocarbon receptor signalling in idiopathic membranous nephropathy. Free Radic Biol Med. 2023;207:89-106
38. Evenepoel P, Stenvinkel P, Shanahan C, Pacifici R. Inflammation and gut dysbiosis as drivers of CKD-MBD. Nat Rev Nephrol. 2023;19:646-57
39. Miao H, Wang YN, Yu XY, Zou L, Guo Y, Su W. et al. Lactobacillus species ameliorate membranous nephropathy through inhibiting the aryl hydrocarbon receptor pathway via tryptophan-produced indole metabolites. Br J Pharmacol. 2024;181:162-79
40. Wang X, Yang S, Li S, Zhao L, Hao Y, Qin J. et al. Aberrant gut microbiota alters host metabolome and impacts renal failure in humans and rodents. Gut. 2020;69:2131-42
41. Zhu H, Cao CJ, Wu ZC, Zhang HP, Sun ZH, Wang M. et al. The probiotic L. casei Zhang slows the progression of acute and chronic kidney disease. Cell Metabolism. 2021;33:1926 -+
42. Correale J, Hohlfeld R, Baranzini SE. The role of the gut microbiota in multiple sclerosis. Nat Rev Neurol. 2022;18:544-58
43. Jin Y, Zhang SJ, Zhuang S, Li P, Miao H, Zhao YY. Microbiota-gut-kidney axis in health and renal disease. Int J Biol Sci. 2026;22:750-70
44. Fan Y, Pedersen O. Gut microbiota in human metabolic health and disease. Nat Rev Microbiol. 2021;19:55-71
45. Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR. et al. Enterotypes of the human gut microbiome. Nature. 2011;473:174-80
46. Tang WH, Kitai T, Hazen SL. Gut microbiota in cardiovascular health and disease. Circ Res. 2017;120:1183-96
47. Hsu CL, Schnabl B. The gut-liver axis and gut microbiota in health and liver disease. Nat Rev Microbiol. 2023;21:719-33
48. Chen YY, Chen DQ, Chen L, Liu JR, Vaziri ND, Guo Y. et al. Microbiome-metabolome reveals the contribution of gut-kidney axis on kidney disease. J Transl Med. 2019;17:5
49. Mann ER, Lam YK, Uhlig HH. Short-chain fatty acids: linking diet, the microbiome and immunity. Nat Rev Immunol. 2024;24:577-95
50. Lavelle A, Sokol H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat Rev Gastroenterol Hepatol. 2020;17:223-37
51. De Vos WM, Nguyen Trung M, Davids M, Liu G, Rios-Morales M, Jessen H. et al. Phytate metabolism is mediated by microbial cross-feeding in the gut microbiota. Nat Microbiol. 2024;9:1812-27
52. Mukhopadhya I, Louis P. Gut microbiota-derived short-chain fatty acids and their role in human health and disease. Nat Rev Microbiol. 2025;23:635-51
53. Nshanian M, Gruber JJ, Geller BS, Chleilat F, Lancaster SM, White SM. et al. Short-chain fatty acid metabolites propionate and butyrate are unique epigenetic regulatory elements linking diet, metabolism and gene expression. Nat Metab. 2025;7:196-211
54. Ikeda T, Nishida A, Yamano M, Kimura I. Short-chain fatty acid receptors and gut microbiota as therapeutic targets in metabolic, immune, and neurological diseases. Pharmacol Ther. 2022;239:108273
55. Liu YJ, Tang B, Wang FC, Tang L, Lei YY, Luo Y. et al. Parthenolide ameliorates colon inflammation through regulating Treg/Th17 balance in a gut microbiota-dependent manner. Theranostics. 2020;10:5225-41
56. Giordano L, Mihaila SM, Eslami Amirabadi H, Masereeuw R. Microphysiological systems to recapitulate the gut-kidney axis. Trends Biotechnol. 2021;39:811-23
57. Qin X, Zhang M, Chen S, Tang Y, Cui J, Ding G. Short-chain fatty acids in fetal development and metabolism. Trends Mol Med. 2025;31:625-39
58. Pinto CJG, Ávila-Gálvez M, Lian Y, Moura-Alves P, Nunes Dos Santos C. Targeting the aryl hydrocarbon receptor by gut phenolic metabolites: A strategy towards gut inflammation. Redox Biol. 2023;61:102622
59. Xue C, Li G, Zheng Q, Gu X, Shi Q, Su Y. et al. Tryptophan metabolism in health and disease. Cell Metab. 2023;35:1304-26
60. Lin X, He K, Gu Z, Zhao X. Emerging chemophysiological diversity of gut microbiota metabolites. Trends Pharmacol Sci. 2024;45:824-38
61. Lim YJ, Sidor NA, Tonial NC, Che A, Urquhart BL. Uremic toxins in the progression of chronic kidney disease and cardiovascular disease: mechanisms and therapeutic targets. Toxins (Basel). 2021;13:142
62. Jia Y, He M, Wang F, Zhan Y, Deng Q, Shen J. et al. Indole-3-lactic acid protects the gut vascular barrier following intestinal ischemia injury through AhR/Nrf2/STAT3 mediated claudin 2 downregulation. Cell Commun Signal. 2025;23:447
63. Liu L, Yu QQ, Zhang YL, Zhou JT, Jin Y, Jiang CH. et al. Renal fibrosis is induced by hyperactive Wnt/β-catenin pathway via microbial-mediated tryptophan metabolism-driven AhR signaling in rodents and humans. Cell Mol Life Sci. 2026;83:182
64. Hayase E, Jenq RR. Too much TMAO and GVHD. Blood. 2020;136:383-5
65. Jiang S, Shui Y, Cui Y, Tang C, Wang X, Qiu X. et al. Gut microbiota dependent trimethylamine N-oxide aggravates angiotensin II-induced hypertension. Redox Biol. 2021;46:102115
66. Buffa JA, Romano KA, Copeland MF, Cody DB, Zhu W, Galvez R. et al. The microbial gbu gene cluster links cardiovascular disease risk associated with red meat consumption to microbiota L-carnitine catabolism. Nat Microbiol. 2022;7:73-86
67. Ravid JD, Kamel MH, Chitalia VC. Uraemic solutes as therapeutic targets in CKD-associated cardiovascular disease. Nat Rev Nephrol. 2021;17:402-16
68. Saaoud F, Liu L, Xu K, Cueto R, Shao Y, Lu Y. et al. Aorta- and liver-generated TMAO enhances trained immunity for increased inflammation via ER stress/mitochondrial ROS/glycolysis pathways. JCI Insight. 2023;8:e158183
69. Mosterd CM, Kanbay M, van den Born BJH, van Raalte DH, Rampanelli E. Intestinal microbiota and diabetic kidney diseases: the role of microbiota and derived metabolites inmodulation of renal inflammation and disease progression. Best Pract Res Clin Endocrinol Metab. 2021;35:101484
70. Spicher P, Brazier F, Laville SM, Liabeuf S, Kamel S, Culot M. et al. Transporter-mediated interactions between uremic toxins and drugs: a hidden driver of toxicity in chronic kidney disease. Int J Mol Sci. 2025;26:6328
71. Peluzio MDG, Martinez JA, Milagro FI. Postbiotics: metabolites and mechanisms involved in microbiota-host interactions. Trends Food Sci Tech. 2021;108:11-26
72. Zhang LS, Davies SS. Microbial metabolism of dietary components to bioactive metabolites: opportunities for new therapeutic interventions. Genome Med. 2016;8:46
73. Nair AV, Singh A, Chakravortty D. Defence Warriors: exploring the crosstalk between polyamines and oxidative stress during microbial pathogenesis. Redox Biol. 2025;83:103648
74. Watanabe S, Nakayama M, Yokoo T, Sanada S, Ubara Y, Komatsuda A. et al. Lubiprostone in chronic kidney disease: Insights into mitochondrial function and polyamines from a randomized phase 2 clinical trial. Sci Adv. 2025;11:eadw3934
75. Nakamura A, Kurihara S, Takahashi D, Ohashi W, Nakamura Y, Kimura S. et al. Symbiotic polyamine metabolism regulates epithelial proliferation and macrophage differentiation in the colon. Nat Commun. 2021;12:2105
76. Fan L, Liu B, Wang Y, Tang B, Xu T, Fu J. et al. Intestinal Lactobacillus murinus-derived small RNAs target porcine polyamine metabolism. Proc Natl Acad Sci U S A. 2024;121:e2413241121
77. Gobert AP, Latour YL, Asim M, Barry DP, Allaman MM, Finley JL. et al. Protective role of spermidine in colitis and colon carcinogenesis. Gastroenterology. 2022;162:813-27.e8
78. Zarbock A, Forni LG, Ostermann M, Ronco C, Bagshaw SM, Mehta RL. et al. Designing acute kidney injury clinical trials. Nat Rev Nephrol. 2024;20:137-46
79. Bejoy J, Qian ES, Woodard LE. Tissue culture models of AKI: from tubule cells to human kidney organoids. J Am Soc Nephrol. 2022;33:487-501
80. Kellum JA, Romagnani P, Ashuntantang G, Ronco C, Zarbock A, Anders HJ. Acute kidney injury. Nat Rev Dis Primers. 2021;7:52
81. Li XJ, Suo P, Wang YN, Zou L, Nie XL, Zhao YY. et al. Arachidonic acid metabolism as a therapeutic target in AKI-to-CKD transition. Front Pharmacol. 2024;15:1365802
82. Suzuki T. Regulation of intestinal epithelial permeability by tight junctions. Cell Mol Life Sci. 2013;70:631-59
83. Zhang J, Ankawi G, Sun J, Digvijay K, Yin Y, Rosner MH. et al. Gut-kidney crosstalk in septic acute kidney injury. Crit Care. 2018;22:117
84. Ruan Q, Lu H, Zhu H, Guo Y, Bai Y. A network-regulative pattern in the pathogenesis of kidney injury following severe acute pancreatitis. Biomed Pharmacother. 2020;125:109978
85. Francis A, Harhay MN, Ong ACM, Tummalapalli SL, Ortiz A, Fogo AB. et al. Chronic kidney disease and the global public health agenda: an international consensus. Nat Rev Nephrol. 2024;20:473-85
86. Ndongo M, Nehemie LM, Coundoul B, Diouara AAM, Seck SM. Prevalence and outcomes of polycystic kidney disease in African populations: a systematic review. World J Nephrol. 2024;13:90402
87. Khandpur S, Mishra P, Mishra S, Tiwari S. Challenges in predictive modelling of chronic kidney disease: a narrative review. World J Nephrol. 2024;13:97214
88. Vlahou A, Vanholder R. Urine as a source of biomarkers and biological knowledge in chronic kidney disease. Nat Rev Nephrol. 2025;22:69-84
89. Zhong J, Kirabo A, Yang HC, Fogo AB, Shelton EL, Kon V. Intestinal lymphatic dysfunction in kidney disease. Circ Res. 2023;132:1226-45
90. Poesen R, Ramezani A, Claes K, Augustijns P, Kuypers D, Barrows IR. et al. Associations of soluble CD14 and endotoxin with mortality, cardiovascular disease, and progression of kidney disease among patients with CKD. Clin J Am Soc Nephrol. 2015;10:1525-33
91. Holle J, Bartolomaeus H, Löber U, Behrens F, Bartolomaeus TUP, Anandakumar H. et al. Inflammation in children with CKD linked to gut dysbiosis and metabolite imbalance. J Am Soc Nephrol. 2022;33:2259-75
92. Dodd D, Spitzer MH, Van Treuren W, Merrill BD, Hryckowian AJ, Higginbottom SK. et al. A gut bacterial pathway metabolizes aromatic amino acids into nine circulating metabolites. Nature. 2017;551:648-52
93. Yang J, Kim CJ, Go YS, Lee HY, Kim MG, Oh SW. et al. Intestinal microbiota control acute kidney injury severity by immune modulation. Kidney Int. 2020;98:932-46
94. Oh CJ, Kim MJ, Lee JM, Kim DH, Kim IY, Park S. et al. Inhibition of pyruvate dehydrogenase kinase 4 ameliorates kidney ischemia-reperfusion injury by reducing succinate accumulation during ischemia and preserving mitochondrial function during reperfusion. Kidney Int. 2023;104:724-39
95. Ni J, Yang Z, Sun X, Cui Q, Zhang R, Lu H. et al. Probiotic Akkermansia muciniphila alleviates acute kidney injury by protecting the intestinal barrier and modulating gut microbiota and metabolites. J Biomed Res. 2025;40:76-88
96. Nakade Y, Iwata Y, Furuichi K, Mita M, Hamase K, Konno R. et al. Gut microbiota-derived D-serine protects against acute kidney injury. JCI Insight. 2018;3:e97957
97. Jin S, Wijerathne CUB, Siow YL, O K. Disruption of the transsulfuration pathway by acute kidney injury causes intestinal damage. iScience. 2025;28:113511
98. He XF, Guo Y, Zhao YY. Targeting gut microbiota as a promising therapy for chronic kidney disease. Integrative Medicine in Nephrology and Andrology. 2025;12:e25-00026
99. Laiola M, Koppe L, Larabi A, Thirion F, Lange C, Quinquis B. et al. Toxic microbiome and progression of chronic kidney disease: insights from a longitudinal CKD-Microbiome Study. Gut. 2025;74:1624-37
100. Ma L, Zhang L, Li J, Zhang X, Xie Y, Li X. et al. The potential mechanism of gut microbiota-microbial metabolites-mitochondrial axis in progression of diabetic kidney disease. Mol Med. 2023;29:148
101. Linh HT, Iwata Y, Senda Y, Sakai-Takemori Y, Nakade Y, Oshima M. et al. Intestinal bacterial translocation contributes to diabetic kidney disease. J Am Soc Nephrol. 2022;33:1105-19
102. Zhao J, Bai M, Ning X, Qin Y, Wang Y, Yu Z. et al. Expansion of Escherichia-Shigella in gut is associated with the onset and response to immunosuppressive therapy of IgA nephropathy. J Am Soc Nephrol. 2022;33:2276-92
103. Tan J, Dong L, Jiang Z, Tan L, Luo X, Pei G. et al. Probiotics ameliorate IgA nephropathy by improving gut dysbiosis and blunting NLRP3 signaling. J Transl Med. 2022;20:382
104. Shang J, Zhang Y, Guo R, Liu W, Zhang J, Yan G. et al. Gut microbiome analysis can be used as a noninvasive diagnostic tool and plays an essential role in the onset of membranous nephropathy. Adv Sci (Weinh). 2022;9:e2201581
105. Tao P, Huo J, Chen L. Bibliometric analysis of the relationship between gut microbiota and chronic kidney disease from 2001-2022. Integr Med Nephrol Androl. 2024;11:e00017
106. Ren Z, Fan Y, Li A, Shen Q, Wu J, Ren L. et al. Alterations of the human gut microbiome in chronic kidney disease. Adv Sci (Weinh). 2020;7:2001936
107. Wu IW, Lin CY, Chang LC, Lee CC, Chiu CY, Hsu HJ. et al. Gut microbiota as diagnostic tools for mirroring disease progression and circulating nephrotoxin levels in chronic kidney disease: discovery and validation study. Int J Biol Sci. 2020;16:420-34
108. Li N, Wang Y, Zhang H, Ma L, Huang X, Qiao Y. et al. Urobiome of patients with diabetic kidney disease in different stages is revealed by 2bRAD-M. J Transl Med. 2025;23:414
109. Koppe L, Soulage CO. The impact of dietary nutrient intake on gut microbiota in the progression and complications of chronic kidney disease. Kidney Int. 2022;102:728-39
110. Andrade-Oliveira V, Amano MT, Correa-Costa M, Castoldi A, Felizardo RJ, de Almeida DC. et al. Gut Bacteria Products Prevent AKI Induced by Ischemia-Reperfusion. J Am Soc Nephrol. 2015;26:1877-88
111. Li HB, Xu ML, Xu XD, Tang YY, Jiang HL, Li L. et al. Faecalibacterium prausnitzii attenuates CKD via butyrate-renal GPR43 Axis. Circ Res. 2022;131:e120-e34
112. Wu IW, Liao YC, Tsai TH, Lin CH, Shen ZQ, Chan YH. et al. Machine-learning assisted discovery unveils novel interplay between gut microbiota and host metabolic disturbance in diabetic kidney disease. Gut Microbes. 2025;17:2473506
113. Li YJ, Chen X, Kwan TK, Loh YW, Singer J, Liu Y. et al. Dietary fiber protects against diabetic nephropathy through short-chain fatty acid-mediated activation of G protein-coupled receptors GPR43 and GPR109A. J Am Soc Nephrol. 2020;31:1267-81
114. Tain YL, Hou CY, Chang-Chien GP, Lin SF, Hsu CN. Perinatal propionate supplementation protects adult male offspring from maternal chronic kidney disease-induced hypertension. Nutrients. 2022;14:3435
115. Corte-Iglesias V, Saiz ML, Andrade-Lopez AC, Salazar N, Bernet CR, Martin-Martin C. et al. Propionate and butyrate counteract renal damage and progression to chronic kidney disease. Nephrol Dial Transplant. 2024;40:133-50
116. Tian X, Zeng Y, Tu Q, Jiao Y, Yao S, Chen Y. et al. Butyrate alleviates renal fibrosis in CKD by regulating NLRP3-mediated pyroptosis via the STING/NF-κB/p65 pathway. Int Immunopharmacol. 2023;124:111010
117. Huang W, Man Y, Gao C, Zhou L, Gu J, Xu H. et al. Short-chain fatty acids ameliorate diabetic nephropathy via GPR43-mediated inhibition of oxidative stress and NF-κB signaling. Oxid Med Cell Longev. 2020;2020:4074832
118. Chen Q, Liang X, Wu T, Jiang J, Jiang Y, Zhang S. et al. Integrative analysis of metabolomics and proteomics reveals amino acid metabolism disorder in sepsis. J Transl Med. 2022;20:123
119. Walker JA, Richards S, Belghasem ME, Arinze N, Yoo SB, Tashjian JY. et al. Temporal and tissue-specific activation of aryl hydrocarbon receptor in discrete mouse models of kidney disease. Kidney Int. 2020;97:538-50
120. Miao Y, Wang M, Sun H, Zhang Y, Zhou W, Yang W. et al. Bifidobacterium longum metabolite indole-3-carboxaldehyde blocks HDAC3 and inhibits macrophage NLRP3 inflammasome activation in intestinal ischemia/reperfusion injury. Inflammation. 2025;48:2572-87
121. Balestrieri N, Palzkill V, Pass C, Tan J, Salyers ZR, Moparthy C. et al. Activation of the aryl hydrocarbon receptor in muscle exacerbates ischemic pathology in chronic kidney disease. Circ Res. 2023;133:158-76
122. Hou Q, Ye L, Liu H, Huang L, Yang Q, Turner JR. et al. Lactobacillus accelerates ISCs regeneration to protect the integrity of intestinal mucosa through activation of STAT3 signaling pathway induced by LPLs secretion of IL-22. Cell Death Differ. 2018;25:1657-70
123. Krishnan S, Ding Y, Saedi N, Choi M, Sridharan GV, Sherr DH. et al. Gut microbiota-derived tryptophan metabolites modulate inflammatory response in hepatocytes and macrophages. Cell Rep. 2018;23:1099-111
124. Yu J, Luo Y, Zhu Z, Zhou Y, Sun L, Gao J. et al. A tryptophan metabolite of the skin microbiota attenuates inflammation in patients with atopic dermatitis through the aryl hydrocarbon receptor. J Allergy Clin Immunol. 2019;143:2108-19.e12
125. Corradi V, Caprara C, Barzon E, Mattarollo C, Zanetti F, Ferrari F. et al. A possible role of p-cresyl sulfate and indoxyl sulfate as biomarkers in the prediction of renal function according to the GFR (G) categories. Integr Med Nephrol Androl. 2024;11:e24-00002
126. Kim HY, Yoo TH, Cho JY, Kim HC, Lee WW. Indoxyl sulfate-induced TNF-α is regulated by crosstalk between the aryl hydrocarbon receptor, NF-κB, and SOCS2 in human macrophages. FASEB J. 2019;33:10844-58
127. Dou L, Sallée M, Cerini C, Poitevin S, Gondouin B, Jourde-Chiche N. et al. The cardiovascular effect of the uremic solute indole-3 acetic acid. J Am Soc Nephrol. 2015;26:876-87
128. Vial R, Poitevin S, McKay N, Burtey S, Cerini C. Tryptophan metabolites regulate neuropentraxin 1 expression in endothelial cells. Int J Mol Sci. 2022;23:2369
129. Xie H, Yang N, Lu L, Sun X, Li J, Wang X. et al. Uremic toxin receptor AhR facilitates renal senescence and fibrosis via suppressing mitochondrial biogenesis. Adv Sci (Weinh). 2024;11:e2402066
130. Zhao BR, Hu XR, Wang WD, Zhou Y. Cardiorenal syndrome: clinical diagnosis, molecular mechanisms and therapeutic strategies. Acta Pharmacol Sin. 2025;46:1539-55
131. Lee J, Lee J, Kim K, Lee J, Jung Y, Hyeon JS. et al. Antibiotic-induced intestinal microbiota depletion can attenuate the acute kidney injury to chronic kidney disease transition via NADPH oxidase 2 and trimethylamine-N-oxide inhibition. Kidney Int. 2024;105:1239-53
132. Wang M, Tang WHW, Li XS, de Oliveira Otto MC, Lee Y, Lemaitre RN. et al. The gut microbial metabolite trimethylamine N-oxide, incident CKD, and kidney function decline. J Am Soc Nephrol. 2024;35:749-60
133. Tang WH, Wang Z, Kennedy DJ, Wu Y, Buffa JA, Agatisa-Boyle B. et al. Gut microbiota-dependent trimethylamine N-oxide (TMAO) pathway contributes to both development of renal insufficiency and mortality risk in chronic kidney disease. Circ Res. 2015;116:448-55
134. Andrikopoulos P, Aron-Wisnewsky J, Chakaroun R, Myridakis A, Forslund SK, Nielsen T. et al. Evidence of a causal and modifiable relationship between kidney function and circulating trimethylamine N-oxide. Nat Commun. 2023;14:5843
135. Cheng E, Hung SC, Lin TY. Association of trimethylamine N-oxide and metabolites with kidney function decline in patients with chronic kidney disease. Clin Nutr. 2025;44:239-47
136. Xie Y, Hu X, Li S, Qiu Y, Cao R, Xu C. et al. Pharmacological targeting macrophage phenotype via gut-kidney axis ameliorates renal fibrosis in mice. Pharmacol Res. 2022;178:106161
137. Dong F, Jiang S, Tang C, Wang X, Ren X, Wei Q. et al. Trimethylamine N-oxide promotes hyperoxaluria-induced calcium oxalate deposition and kidney injury by activating autophagy. Free Radic Biol Med. 2022;179:288-300
138. Lee TH, Park D, Kim YJ, Lee I, Kim S, Oh CT. et al. Lactobacillus salivarius BP121 prevents cisplatin-induced acute kidney injury by inhibition of uremic toxins such as indoxyl sulfate and p-cresol sulfate via alleviating dysbiosis. Int J Mol Med. 2020;45:1130-40
139. Chen L, Shi J, Ma X, Shi D, Qu H. Effects of microbiota-driven therapy on circulating indoxyl sulfate and p-cresyl sulfate in patients with chronic kidney disease: a systematic review and meta-analysis of randomized controlled trials. Adv Nutr. 2022;13:1267-78
140. Balint L, Socaciu C, Socaciu AI, Vlad A, Gadalean F, Bob F. et al. Quantitative, Targeted Analysis of Gut Microbiota Derived Metabolites Provides Novel Biomarkers of Early Diabetic Kidney Disease in Type 2 Diabetes Mellitus Patients. Biomolecules. 2023;13:1086
141. Freise C, Metzkow S, Zappe A, Ebert M, Stolzenburg N, Hahndorf J. et al. The uremic toxins inorganic phosphate, indoxylsulphate, p-cresylsulphate, and TMAO induce the generation of sulphated glycosaminoglycans in aortic tissue and vascular cells via pAKT signaling: a missing link in the "gut-matrix axis". Toxins (Basel). 2025;17:217
142. Watanabe H, Miyamoto Y, Honda D, Tanaka H, Wu Q, Endo M. et al. P-Cresyl sulfate causes renal tubular cell damage by inducing oxidative stress by activation of NADPH oxidase. Kidney Int. 2013;83:582-92
143. Wu IW, Gao SS, Chou HC, Yang HY, Chang LC, Kuo YL. et al. Integrative metagenomic and metabolomic analyses reveal severity-specific signatures of gut microbiota in chronic kidney disease. Theranostics. 2020;10:5398-411
144. Li X, Zhou X, Liu X, Li X, Jiang X, Shi B. et al. Spermidine protects against acute kidney injury by modulating macrophage NLRP3 inflammasome activation and mitochondrial respiration in an eIF5A hypusination-related pathway. Mol Med. 2022;28:103
145. Feng YL, Cao G, Chen DQ, Vaziri ND, Chen L, Zhang J. et al. Microbiome-metabolomics reveals gut microbiota associated with glycine-conjugated metabolites and polyamine metabolism in chronic kidney disease. Cell Mol Life Sci. 2019;76:4961-78
146. Nishiyama K, Aono K, Fujimoto Y, Kuwamura M, Okada T, Tokumoto H. et al. Chronic kidney disease after 5/6 nephrectomy disturbs the intestinal microbiota and alters intestinal motility. J Cell Physiol. 2019;234:6667-78
147. Zahedi K, Barone S, Brooks M, Stewart TM, Foley JR, Nwafor A. et al. Polyamine catabolism and its role in renal injury and fibrosis in mice subjected to repeated low-dose cisplatin treatment. Biomedicines. 2024;12:640
148. Aihara S, Torisu K, Uchida Y, Imazu N, Nakano T, Kitazono T. Spermidine from arginine metabolism activates Nrf2 and inhibits kidney fibrosis. Commun Biol. 2023;6:676
149. Luo D, Lu X, Li H, Li Y, Wang Y, Jiang S. et al. The spermine oxidase/spermine axis coordinates ATG5-mediated autophagy to orchestrate renal senescence and fibrosis. Adv Sci (Weinh). 2024;11:e2306912
150. Zixin Y, Lulu C, Xiangchang Z, Qing F, Binjie Z, Chunyang L. et al. TMAO as a potential biomarker and therapeutic target for chronic kidney disease: A review. Front Pharmacol. 2022;13:929262
151. Barrios C, Beaumont M, Pallister T, Villar J, Goodrich JK, Clark A. et al. Gut-microbiota-metabolite axis in early renal function decline. PLoS One. 2015;10:e0134311
152. Atzeni A, Díaz-López A, Cacho AH, Babio N, García-Gavilán JF, Cornejo-Pareja I. et al. Gut microbiota dynamics and association with chronic kidney disease: A longitudinal study within the PREDIMED-Plus trial. Life Sci. 2024;351:122863
153. Szeto CC, Ng JKC. Toxic microbiome and progression of chronic kidney disease: insights from a longitudinal CKD-microbiome study. Gut. 2025;2025:335600
154. Stepanova N. Probiotic interventions in peritoneal dialysis: a review of underlying mechanisms and therapeutic potentials. World J Nephrol. 2024;13:98719
155. Lin TY, Wu PH, Lin YT, Hung SC. Gut dysbiosis and mortality in hemodialysis patients. NPJ Biofilms Microbiomes. 2021;7:20
156. Basile G, Pecoraro A, Gallioli A, Territo A, Berquin C, Robalino J. et al. Robotic kidney transplantation. Nat Rev Urol. 2024;21:521-33
157. Hunter J, Hosgood S, Moers C, Leuvenink H, Rabelink TJ, Ploeg R. Improving outcomes in kidney transplantation through advances in donor organ perfusion. Nat Rev Nephrol. 2025;21:818-32
158. Shdaifat EA, Abu-Sneineh FT, Ibrahim AM. Quality of life and psychological distress in end-stage renal disease patients undergoing hemodialysis and transplantation. World J Nephrol. 2024;13:95739
159. Zhang L, Xie F, Tang H, Zhang X, Hu J, Zhong X. et al. Gut microbial metabolite TMAO increases peritoneal inflammation and peritonitis risk in peritoneal dialysis patients. Transl Res. 2022;240:50-63
160. McIntyre CW, Jain A. Dialysis and cognitive impairment. Nat Rev Nephrol. 2025;21:553-64
161. Raikou VD. Renoprotective strategies. World J Nephrol. 2024;13:89637
162. Agarwal AK, Haddad NJ, Vachharajani TJ, Asif A. Innovations in vascular access for hemodialysis. Kidney Int. 2019;95:1053-63
163. Bello AK, Okpechi IG, Osman MA, Cho Y, Cullis B, Htay H. et al. Epidemiology of peritoneal dialysis outcomes. Nat Rev Nephrol. 2022;18:779-93
164. Meyer TW, Bargman JM. The removal of uremic solutes by peritoneal dialysis. J Am Soc Nephrol. 2023;34:1919-27
165. Du J, Zhao X, Ding X, Ren Q, Wang H, Han Q. et al. Gut microbiota and their metabolites in hemodialysis patients. Chin Med J (Engl). 2025;138:502-4
166. Mahdavi S, Anthony NM, Sikaneta T, Tam PY. Perspective: multiomics and artificial intelligence for personalized nutritional management of diabetes in patients undergoing peritoneal dialysis. Adv Nutr. 2025;16:100378
167. Li Y, Dai M, Yan J, Liu F, Wang X, Lin L. et al. Colonic dialysis can influence gut flora to protect renal function in patients with pre-dialysis chronic kidney disease. Sci Rep. 2021;11:12773
168. Dai S, Dai Y, Peng J, Xie X, Ning J. Simplified colonic dialysis with hemodialysis solutions delays the progression of chronic kidney disease. Qjm. 2019;112:189-96
169. Kajbafzadeh AM, Zeinoddini A, Heidari R, NaserHodjjati H, Tourchi A. A novel alternative for renal replacement therapy: 2-year successful colonic dialysis via a Malone antegrade continent enema stoma. J Pediatr Urol. 2014;10:511-4
170. Kajbafzadeh AM, Sabetkish N, Sabetkish S. Establishment of colonic dialysis model in uremic rats by right nephrectomy and left partial nephrectomy. J Pediatr Urol. 2018;14:159.e1-e8
171. Wang Z, Zhang S, Zheng X, Zhang L. Efficacy and safety of colonic dialysis combined with traditional Chinese medicine retention enema in the treatment of chronic renal failure: A protocol for systematic review and meta-analysis. Medicine (Baltimore). 2021;100:e28082
172. He H, Shen M, Sun W, Tang Y, Xu X, Xie Y. Influence of colonic dialysis using Chinese medicine on creatinine decomposition by intestinal bacteria in uremia rats. Am J Transl Res. 2019;11:6577-84
173. He H, Hu P, Tang Y, Xu X. Influence of colonic dialysis using Gubenxiezhuo on the distribution of gut microflora in uremia rats. J Cell Physiol. 2019;234:11882-7
174. Mikami N, Sakaguchi S. Regulatory T cells in autoimmune kidney diseases and transplantation. Nat Rev Nephrol. 2023;19:544-57
175. Tasca P, van den Berg BM, Rabelink TJ, Wang G, Heijs B, van Kooten C. et al. Application of spatial-omics to the classification of kidney biopsy samples in transplantation. Nat Rev Nephrol. 2024;20:755-66
176. Peticca B, Prudencio TM, Robinson SG, Karhadkar SS. Challenges with non-descriptive compliance labeling of end-stage renal disease patients in accessibility for renal transplantation. World J Nephrol. 2024;13:88967
177. Gadelkareem RA, Abdelgawad AM, Reda A, Azoz NM, Zarzour MA, Mohammed N. et al. Preemptive living donor kidney transplantation: access, fate, and review of the status in Egypt. World J Nephrol. 2023;12:40-55
178. Swarte JC, Zhang S, Nieuwenhuis LM, Gacesa R, Knobbe TJ, De Meijer VE. et al. Multiple indicators of gut dysbiosis predict all-cause and cause-specific mortality in solid organ transplant recipients. Gut. 2024;73:1650-61
179. Arinze NV, Yin W, Lotfollahzadeh S, Napoleon MA, Richards S, Walker JA. et al. Tryptophan metabolites suppress the Wnt pathway and promote adverse limb events in chronic kidney disease. J Clin Invest. 2022;132:e142260
180. Yepes-Calderón M, Martín Del Campo Sanchez F, Kremer D, Knobbe TJ, Gomes Neto AW, Connelley MA. et al. Plasma trimethylamine N-oxide concentration and all-cause mortality in kidney transplant recipients. Nephrol Dial Transplant. 2025;40:1931-40
181. Wu H, Singer J, Kwan TK, Loh YW, Wang C, Tan J. et al. Gut microbial metabolites induce donor-specific tolerance of kidney allografts through induction of T regulatory cells by short-chain fatty acids. J Am Soc Nephrol. 2020;31:1445-61
182. Cai R, Li C, Zhao Y, Yuan H, Zhang X, Liang A. et al. Traditional Chinese medicine in diabetic kidney disease: multifaceted therapeutic mechanisms and research progress. Chin Med. 2025;20:95
183. Acquah-Mills J, Henneh IT, Arhin SM, Ekor M. The prevalence, pattern of use and impact of traditional, complementary and alternative medicine (TCAM) among patients with end stage renal disease undergoing haemodialysis at a tertiary health facility in Ghana. Clin Tradit Med Pharmacol. 2025;6:200232
184. Guo ZY, Wu X, Zhang SJ, Yang JH, Miao H, Zhao YY. Poria cocos: traditional uses, triterpenoid components and their renoprotective pharmacology. Acta Pharmacol Sin. 2025;46:836-51
185. Feng HY, Wang YQ, Yang J, Miao H, Zhao YY, Li X. Anthraquinones from Rheum officinale ameliorate renal fibrosis in acute kidney injury and chronic kidney disease. Drug Des Devel Ther. 2025;19:5739-60
186. Wu X, Sun W, Xing L, Zhao J. Effect of Yishenqinglihuoxue formula in the treatment of patients with stage 5 chronic kidney disease using propensity score matching: a retrospective clinical study. Clin Tradit Med Pharmacol. 2025;6:200239
187. Wang YN, Li XJ, Wang WF, Zou L, Miao H, Zhao YY. Geniposidic acid attenuates chronic tubulointerstitial nephropathy through regulation of the NF-ƙB/Nrf2 pathway via aryl hydrocarbon receptor signaling. Phytother Res. 2024;38:5441-57
188. Wang W, Dai R, Cheng M, Chen Y, Gao Y, Hong X. et al. Metabolic reprogramming and renal fibrosis: what role might Chinese medicine play? Chin Med. 2024;19:148
189. Gao X, Chen L, Zhang C, Gao H, Yang S, Huang G. et al. Total Panax notoginseng saponins treats diabetic kidney disease in mice by regulating the TXNIP/NLRP3 signaling pathway. Clin Tradit Med Pharmacol. 2025;6:200245
190. Miao H, Wang KE, Li P, Zhao YY. Rhubarb: traditional uses, phytochemistry, multiomics-based novel pharmacological and toxicological mechanisms. Drug Des Devel Ther. 2025;19:9457-80
191. Shao YF, Tang BB, Ding YH, Fang CY, Hong L, Shao CX. et al. Kaempferide ameliorates cisplatin-induced nephrotoxicity via inhibiting oxidative stress and inducing autophagy. Acta Pharmacol Sin. 2023;44:1442-54
192. Liu J, Vaziri ND, Miao H, Zhao YY. The renoprotective effect, nephrotoxicity, and molecular mechanisms of emodin. Integr Med Nephrol Androl. 2026;13:e25-00036
193. Kuang BC, Wang ZH, Hou SH, Zhang J, Wang MQ, Zhang JS. et al. Methyl eugenol protects the kidney from oxidative damage in mice by blocking the Nrf2 nuclear export signal through activation of the AMPK/GSK3β axis. Acta Pharmacol Sin. 2023;44:367-80
194. Liang BE, Long LS, Wu XY, Huang MY, Lai Y, Yuan X. et al. Alginate oligosaccharide prevents renal ischemia-reperfusion injury in rats via MRC1-mediated pathway. Acta Pharmacol Sin. 2025;46:2450-67
195. Pan D, Qu Y, Shi C, Xu C, Zhang J, Du H. et al. Oleanolic acid and its analogues: promising therapeutics for kidney disease. Chin Med. 2024;19:74
196. Yang X, Xin YJ, Gu YZ, Wang YL, Hu XJ, Ying GH. et al. Total alkaloids of Aconitum carmichaelii Debx alleviate cisplatin-induced acute renal injury by inhibiting inflammation and oxidative stress related to gut microbiota metabolism. Phytomedicine. 2024;135:156128
197. Li J, Tan YJ, Wang MZ, Sun Y, Li GY, Wang QL. et al. Loganetin protects against rhabdomyolysis-induced acute kidney injury by modulating the toll-like receptor 4 signalling pathway. Br J Pharmacol. 2019;176:1106-21
198. Mo P, Gao B, Wang R, Huang S, Chen Q, Li M. et al. Knowledge mapping and global research trends of ginseng polysaccharides: a bibliometric analysis with visualizations from 1985 to 2023. Drug Des Devel Ther. 2025;19:2749-66
199. Nie YM, Zhou WQ, Niu T, Mao MF, Zhan YX, Li Y. et al. Peptidoglycan isolated from the fruit of Lycium barbarum alleviates liver fibrosis in mice by regulating the TGF-β/Smad7 signaling and gut microbiota. Acta Pharmacol Sin. 2025;46:1329-44
200. Zhang QW, Yang MJ, Liao CY, Taha R, Li QY, Abdelmotalab MI. et al. Atractylodes macrocephala Koidz polysaccharide ameliorates DSS-induced colitis in mice by regulating the gut microbiota and tryptophan metabolism. Br J Pharmacol. 2025;182:1508-27
201. Lu X, Guo C, Zhu Y. Selenium-enriched crude polysaccharide from Rosa roxburghii Tratt ameliorates cadmium-induced acute kidney injury in mice by modulating intestinal microorganisms. Heliyon. 2023;9:e19678
202. Zhang YL, Feng XE, Chang RR, Zhang LH, Xiao BG, Li QS. et al. Therapeutic effects of 5,2'-dibromo-2,4',5'-trihydroxydiphenylmethanone (LM49) in an experimental rat model of acute pyelonephritis by immunomodulation and anti-inflammation. Int Immunopharmacol. 2018;62:155-64
203. Wang YN, Wu X, Shan QY, Yang Q, Yu XY, Yang JH. et al. Acteoside-containing caffeic acid is bioactive functional group of antifibrotic effect by suppressing inflammation via inhibiting AHR nuclear translocation in chronic kidney disease. Acta Pharmacol Sin. 2025;46:2975-88
204. Wang B, Yang LN, Yang LT, Liang Y, Guo F, Fu P. et al. Fisetin ameliorates fibrotic kidney disease in mice via inhibiting ACSL4-mediated tubular ferroptosis. Acta Pharmacol Sin. 2024;45:150-65
205. Ajibade TO, Oliyide EO, Esan OO, Adetona MO, Awoyomi OV, Oyagbemi TO. et al. Protective effects of naringin on fipronil-induced cardiovascular and renal dysfunctions in rats. Clin Tradit Med Pharmacol. 2024;5:200138
206. Li XJ, Wang YN, Wang WF, Nie X, Miao H, Zhao YY. Barleriside A, an aryl hydrocarbon receptor antagonist, ameliorates podocyte injury through inhibiting oxidative stress and inflammation. Front Pharmacol. 2024;15:1386604
207. Miao H, Vaziri ND. Hederagenin: molecular mechanisms and therapeutic potential for chronic kidney disease. Integr Med Nephrol Androl. 2025;12:e25-00017
208. Ben-Azu B, Asiwe JN, Oritsemuelebi B, Chidebe EO, Onuelu JE, Isibor H. et al. Morin inhibits oxidative stress and inflammation in the cardiorenal system associated with post-traumatic stress and alcohol use disorders in mice. Clin Tradit Med Pharmacol. 2025;6:200196
209. Zheng X, Wang L, Cheng Y, Lin H, Liu S, Chen X. et al. Rhein alleviates renal interstitial fibrosis by inhibiting Smad3 phosphorylation in TGF-β/Smad signalling pathway. Chin Herb Med. 2025;17:744-55
210. Dong X, Zhang J, Li W, Li Y, Jia L, Liu Z. et al. Yi-Shen-Hua-Shi regulates intestinal microbiota dysbiosis and protects against proteinuria in patients with chronic kidney disease: a randomized controlled study. Pharm Biol. 2024;62:356-66
211. Liu J, Gao LD, Fu B, Yang HT, Zhang L, Che SQ. et al. Efficacy and safety of Zicuiyin decoction on diabetic kidney disease: a multicenter, randomized controlled trial. Phytomedicine. 2022;100:154079
212. Lin W, Jiang C, Yu H, Wang L, Li J, Liu X. et al. The effects of Fushen Granule on the composition and function of the gut microbiota during peritoneal dialysis-related peritonitis. Phytomedicine. 2021;86:153561
213. Jiang C-H, Zhang S-J, Li P, Miao H, Zhao Y-Y. Natural products targeting TGF-β/Smad signaling in renal fibrosis: multiomics-based novel molecular mechanisms and therapeutic strategies. Phytomedicine. 2025: 157496.
214. Xin L, Jing X, Li H, Wenfan G, Ming C. Efficacy and safety of activating blood circulation and removing blood stasis of Traditional Chinese Medicine for managing renal fibrosis in patients with chronic kidney disease: a systematic review and Meta-analysis. J Tradit Chin Med. 2023;43:429-40
215. Zhang L, Miao R, Yu T, Wei R, Tian F, Huang Y. et al. Comparative effectiveness of traditional Chinese medicine and angiotensin converting enzyme inhibitors, angiotensin receptor blockers, and sodium glucose cotransporter inhibitors in patients with diabetic kidney disease: A systematic review and network meta-analysis. Pharmacol Res. 2022;177:106111
216. Zhao J, Tostivint I, Xu L, Huang J, Gambotti L, Boffa JJ. et al. Efficacy of combined Abelmoschus manihot and irbesartan for reduction of albuminuria in patients with type 2 diabetes and diabetic kidney disease: a multicenter randomized double-blind parallel controlled clinical trial. Diabetes Care. 2022;45:e113-e5
217. Zhang CY, Yue DJ, Wang D, Wu FF. Effects of Bifidobacterium bifidum tetragonum tablets and Jin Gui Ren Qi Pill on intestinal flora and metabolism in patients with diabetic kidney disease. Front Pharmacol. 2024;15:1346168
218. Hu Z, Jia J, Su Y, Gu Y, Cheng B, Jiang N. Modified Zexie decoction improves phlegm-dampness type stage I hypertension by regulating the gut-immune-kidney axis. Front Pharmacol. 2025;16:1578815
219. Chen X, Gao S, Ruan M, Chen S, Xu J, Xing X. et al. Shen-Shuai-Ning granule decreased serum concentrations of indoxyl sulphate in uremic patients undergoing peritoneal dialysis. Biosci Rep. 2018;38:BSR20171694
220. Ren Q, Cheng L, Guo F, Tao S, Zhang C, Ma L. et al. Fisetin improves hyperuricemia-induced chronic kidney disease via regulating gut microbiota-mediated tryptophan metabolism and aryl hydrocarbon receptor activation. J Agric Food Chem. 2021;69:10932-42
221. Pan L, Yu H, Fu J, Hu J, Xu H, Zhang Z. et al. Berberine ameliorates chronic kidney disease through inhibiting the production of gut-derived uremic toxins in the gut microbiota. Acta Pharm Sin B. 2023;13:1537-53
222. Hua Q, Han Y, Zhao H, Zhang H, Yan B, Pei S. et al. Punicalagin alleviates renal injury via the gut-kidney axis in high-fat diet-induced diabetic mice. Food Funct. 2022;13:867-79
223. Pan S, Jiang SS, Li R, Tian B, Huang CY, Wang R. et al. Hong Guo Ginseng Guo (HGGG) protects against kidney injury in diabetic nephropathy by inhibiting NLRP3 inflammasome and regulating intestinal flora. Phytomedicine. 2024;132:155861
224. Si H, Chen Y, Hu D, Yao S, Yang J, Wen X. A graminan type fructan from Achyranthes bidentata prevents the kidney injury in diabetic mice by regulating gut microbiota. Carbohydr Polym. 2024;339:122275
225. Liao H, Zhao Y, Liang Y, Zou K. Flavonoids derived from Opuntia ficus-indica fruit alleviate renal injury in diabetic nephropathy mice by altering gut microbiota and promoting the production of SCFAs. Nutrients. 2025;17:1800
226. Ma Y, Peng L, Song H, Gu J, Sheng Q, Geng B. et al. Polygonum cuspidatum alleviates hyperuricemia-induced renal injury via rectifying amino acid metabolism disorders. Phytomedicine. 2026;150:157653
227. Meng Y, Hou Y, Zhang R, Guo Z, Zhang Z, Li J. et al. Jinlida ameliorates diabetic kidney disease via gut microbiota-dependent production of pyridoxamine targeting renal AGEs/RAGE and TGF-β pathways. Phytomedicine. 2026;150:157744
228. Xu B, Zheng J, Tian X, Yuan F, Liu Z, Zhou Y. et al. Protective mechanism of traditional Chinese medicine guizhi fuling pills against carbon tetrachloride-induced kidney damage is through inhibiting oxidative stress, inflammation and regulating the intestinal flora. Phytomedicine. 2022;101:154129
229. Zhou X, Liu X, Peng B, Yang Y, Lu H, Li D. et al. Simiao Decoction alleviates hyperuricemia-induced renal injury through regulating gut dysbiosis and decreasing gut-derived uremic toxins. NPJ Biofilms Microbiomes. 2026;12:58
230. Huang RZ, Zhang J, Xu XX, Sun MZ, Xu LG, Kuang H. et al. The multiple benefits of bioactive polysaccharides: from the gut to overall health. Trends Food Sci Tech. 2024;152:104677
231. Yu WC, Liu JR, Baranenko D, Cifuentes A, Ibañez E, Zhang YC. et al. The role of dietary polysaccharides in uric acid regulation: mechanisms and benefits in managing hyperuricemia. Trends Food Sci Tech. 2025;157:104902
232. Wang Z, Rong XL, Dai CX, Wang Q, Lu LF, Zhang WT. et al. Fucoidan alleviates renal fibrosis in mice via Akkermansia muciniphila-mediated suppression of NEU1-TLR4-NFκB signaling axis. Phytomedicine. 2025;145:157060
233. Wang Z, Wu G, Niu T, Guo Y, Wang C, Wang X. et al. Polysaccharide isolated from Dioscorea septemloba improves hyperuricemia and alleviates renal fibrosis through gut-kidney axis in mice. Int J Biol Macromol. 2024;282:137112
234. Ma Y, Pan S, Yang T, Liao B, Yang M, Fu Y. et al. Multi-omics analyses reveal a rhamnogalacturonan I (RG-I) from Polygonum aviculare ameliorates nephrolithiasis through regulation of the gut-kidney axis. Carbohydr Polym. 2026;378:124918
235. Kalantar-Zadeh K, Jafar TH, Nitsch D, Neuen BL, Perkovic V. Chronic kidney disease. Lancet. 2021;398:786-802
236. Zhang L, Wang F, Wang R, Sun B, Liu PJ. Effects of probiotics, prebiotics, and synbiotics on cardiometabolic risk factors in children and adolescents with overweight or obesity: a systematic review and Bayesian network meta-analysis. Crit Rev Food Sci Nutr. 2025;65:5287-301
237. Sanders ME, Hill C. The microbiome: An actor or stage for the beneficial action of probiotics, prebiotics, synbiotics, and postbiotics? Cell Host Microbe. 2025;33:777-89
238. Zhang Y, Han X, Feng T, Li Z, Yu H, Chen Y. et al. Gut-microbiota-derived indole sulfate promotes heart failure in chronic kidney disease. Cell Host Microbe. 2025;33:1715-30.e5
239. Tungsanga S, Katavetin P, Panpetch W, Udompornpitak K, Saisorn W, Praditpornsilpa K. et al. Lactobacillus rhamnosus L34 attenuates chronic kidney disease progression in a 5/6 nephrectomy mouse model through the excretion of anti-inflammatory molecules. Nephrol Dial Transplant. 2022;37:1429-42
240. Ebrahim Z, Proost S, Tito RY, Raes J, Glorieux G, Moosa MR. et al. The effect of ß-glucan prebiotic on kidney function, uremic toxins and gut microbiome in stage 3 to 5 chronic kidney disease (CKD) predialysis participants: a randomized controlled trial. Nutrients. 2022;14:805
241. Yang J, Hou L, Wang A, Shang L, Jia X, Xu R. et al. Prebiotics improve frailty status in community-dwelling older individuals in a double-blind, randomized, controlled trial. J Clin Invest. 2024;134:e176507
242. Lakshmanan AP, Al Za'abi M, Ali BH, Terranegra A. The influence of the prebiotic gum acacia on the intestinal microbiome composition in rats with experimental chronic kidney disease. Biomed Pharmacother. 2021;133:110992
243. Tian Z, Zhang J, Tan Z, Duan X, Zhang H, Li B. et al. Lactiplantibacillus plantarum MPB-65 combined with epicatechin alleviates hyperuricemia by modulating the gut-kidney axis. Food Funct. 2025;16:6800-16
244. Cooper TE, Khalid R, Chan S, Craig JC, Hawley CM, Howell M. et al. Synbiotics, prebiotics and probiotics for people with chronic kidney disease. Cochrane Database Syst Rev. 2023;10:Cd013631
245. Lee ZY, Lew CCH, Ortiz-Reyes A, Patel JJ, Wong YJ, Loh CTI. et al. Benefits and harm of probiotics and synbiotics in adult critically ill patients. A systematic review and meta-analysis of randomized controlled trials with trial sequential analysis. Clin Nutr. 2023;42:519-31
246. Podadera-Herreros A, Alcala-Diaz JF, Gutierrez-Mariscal FM, Jimenez-Torres J, Cruz-Ares S, Arenas-de Larriva AP. et al. Long-term consumption of a mediterranean diet or a low-fat diet on kidney function in coronary heart disease patients: the CORDIOPREV randomized controlled trial. Clin Nutr. 2022;41:552-9
247. Xia J, Zhang Y, Zhang S, Lu C, Huan H, Guan X. Oat dietary fiber delays the progression of chronic kidney disease in mice by modulating the gut microbiota and reducing uremic toxin levels. J Agric Food Chem. 2024
Author contact
![]() Corresponding authors: Professor Hua Miao, E-mail: mh77edu.cn; Professor Li-Min Liu Ph.D., M.D., Email: liuliminedu.cn; Professor Ying-Yong Zhao Ph.D., M.D., Email: zyyedu.cn.
Corresponding authors: Professor Hua Miao, E-mail: mh77edu.cn; Professor Li-Min Liu Ph.D., M.D., Email: liuliminedu.cn; Professor Ying-Yong Zhao Ph.D., M.D., Email: zyyedu.cn.