Impact Factor ISSN: 1449-2288
- Issue 12; 2026
- Issue 11; 2026
- Issue 10; 2026
- Issue 9; 2026
- Issue 8; 2026
- Volume 22; 2026
- Past Issues
- Advance Articles
- Editorial Board
- Cover Images
- Index & Coverage
- Cover Suggestion
- Special Issues
Introduction
Evolution of insect P450s
Classification and functional...
Structural diversity of insect...
Functional versatility of insect...
Conclusion and future...
Abbreviations
Supplementary Material
Acknowledgements
References

 Global reach, higher impact
Global reach, higher impactInt J Biol Sci 2026; 22(12):6281-6297. doi:10.7150/ijbs.135523 This issue Cite
Review
Complex Roles of Insect Cytochrome P450s in Chemical Adaptation
Qi-Ren Chen1, Timothy W. Moural1, Fang Zhu1,2 ![]()
1. Department of Entomology, Pennsylvania State University, University Park, PA 16802, USA.
2. Huck Institutes of the Life Sciences, Pennsylvania State University, University Park, PA 16802, USA.
Received 2026-4-2; Accepted 2026-5-29; Published 2026-6-10
Abstract

The remarkable success of insects is largely due to their capacity to adapt to various environmental stresses. Central to their ability to cope with chemical challenges, cytochrome P450 monooxygenases (P450s) form a highly diversified superfamily that enables this capacity through extensive functional and evolutionary plasticity. Here, we examine the diversity of insect P450s, including their evolution, classification, and structural features. We bring together recent advances to highlight the intricate and interconnected roles of P450s, which are repeatedly co-opted across diverse mechanisms of chemical adaptation and extend across conventional clan boundaries. By integrating functional, evolutionary, and structural perspectives, we propose a holistic framework in which insect P450s act as cross-mechanism and cross-clan nodes linking detoxification, cuticular penetration, olfaction, and symbiont-mediated chemical adaptation. This framework provides a systems-level perspective on how P450 diversification shapes insect responses to chemically complex environments.
Keywords: cytochrome P450, metabolic detoxification, pesticide resistance, cuticular penetration, symbiont-mediated modulation
Introduction
Insects are continually challenged by a wide variety of chemical compounds arising from both natural and anthropogenic sources, including plant allelochemicals, microbial metabolites, and synthetic insecticides [1-4]. Many of these compounds impose strong selective pressures due to their toxic, deterrent, or growth-inhibitory effects, whereas others function as informational cues that mediate interactions among insects, plants, and microbes [5, 6]. The ability to tolerate, detoxify, and adapt to these chemical compounds is a fundamental determinant of insect survival, ecological specialization, and evolutionary success [7-9]. As insects have radiated into diverse habitats and adopted highly variable feeding strategies, chemical adaptation has become a central theme in insect physiology, biochemistry, ecology, and pest management research [9, 10].
To persist in these chemically complex environments, insects have evolved multiple adaptive strategies, including behavioral avoidance, reduced cuticular penetration, enhanced enzymatic detoxification, symbiont-mediated metabolism, and target-site insensitivity [1, 9, 11-13]. Among detoxification enzymes, cytochrome P450 monooxygenases (P450s) are particularly important. They represent one of the most versatile and influential enzyme families involved in chemical adaptation [2, 14, 15]. P450s are a large superfamily of heme-thiolate enzymes first discovered in 1958 [14]. They are ubiquitous across all domains of life, including bacteria, protists, plants, fungi, animals, and even viruses, and play central roles in the metabolism of endogenous compounds and exogenous xenobiotics [16]. P450 enzymes exhibit exceptional catalytic versatility because their heme iron center can access multiple electronic and spin states, enabling flexible frontier orbital interactions and supporting multiple reaction pathways [17]. Since the first insect cytochrome P450 gene, CYP6A1, was cloned and sequenced from the house fly (Musca domestica), advances in genome sequencing have revealed an unprecedented level of P450 diversity in insects [14]. Genome assemblies are now available for more than 2,600 insect species [18], exposing extensive lineage-specific expansions and functional diversification of P450 genes across the insect tree of life [19]. Insect P450s perform essential endogenous functions throughout all life stages, including the biosynthesis and metabolism of hormones (e.g. ecdysteroids and juvenile hormones), lipids, and cuticular hydrocarbons, as well as pheromone degradation [20, 21]. At the same time, many insect P450s function as environmentally responsive enzymes that detoxify xenobiotics, including plant secondary metabolites and synthetic insecticides [3, 22]. The remarkable functional diversity of insect P450s likely reflects conserved catalytic quantum chemistry combined with an unusually evolvable protein scaffold. This enables repeated co-option for metabolizing diverse xenobiotics and mediating chemical communication [19, 23].
Although many insect P450s have been functionally characterized, the vast majority of identified P450 genes remain functionally unstudied [24]. Consequently, our understanding of how P450s act in concert to mediate complex chemical adaptation processes remains limited. Recently, several excellent publications have documented the expansion of P450 gene families, diverse regulatory mechanisms, and broad metabolic capacities across insect taxa [2, 3, 19, 24-27]. Building on these foundations, here we focus on the diversity of insect P450s, including their evolution, classification, and structural features. We also synthesize recent advances to highlight the complex and interconnected roles of insect cytochrome P450s, which are often co-opted across multiple mechanisms of chemical adaptation and span traditional P450 clans. Specifically, we emphasize four interrelated aspects: (i) roles of P450s in host-plant adaptation and chemical communication, (ii) P450s contributions to insecticide resistance and metabolism, (iii) cuticle-associated P450s in chemical adaptation, and (iv) symbiont-mediated modulation of P450 detoxification. By integrating functional, evolutionary, and structural perspectives, we propose a holistic framework for insect P450s, including hormone-biosynthetic and detoxification-associated families. In this framework, P450s act as cross-mechanism and cross-clan nodes linking detoxification, cuticular penetration, olfaction, and symbiont-mediated chemical adaptation.
Evolution of insect P450s
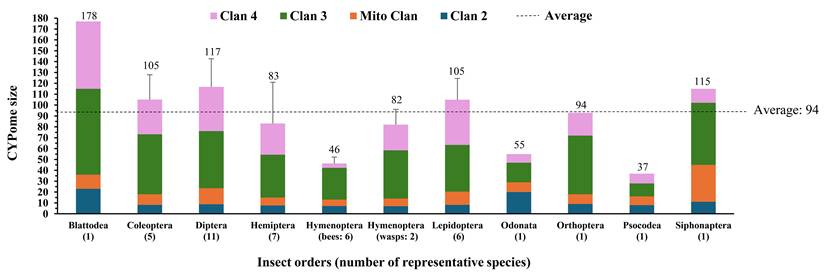
Although the evolutionary origin of the cytochrome P450 superfamily remains unclear, several hypotheses offer insight into the biochemical pressures that have shaped these enzymes [28]. Before acquiring the ability to bind oxygen, early P450s may have functioned as reductases in oxygen-free environments or as peroxygenases catalyzing basic oxidative reactions [29, 30]. With the rise of atmospheric and cellular oxygen, reactive oxygen species (ROS) posed significant threats to cellular integrity. Early P450 enzymes were recruited to detoxify ROS, mitigating oxidative damage to critical biomolecules [31]. The oxygenated products of these reactions, such as sterols and membrane lipids, subsequently assumed important roles as metabolites and signaling molecules, thereby creating new chemical substrates for P450-catalyzed transformations [32, 33]. In insects, this ancient versatility was retained and further elaborated. P450 enzymes perform essential roles in the biosynthesis of hormones, pheromones, and other key metabolites that regulate development, reproduction, and communication [14, 34, 35]. Subsequent ecological pressures, particularly exposure to plant secondary metabolites and other xenobiotics, promoted the expansion and functional diversification of insect P450s, enhancing their capacity for detoxification and chemical adaptation. Host plant chemistry and dietary breadth exert strong selective pressure on these genes, with generalist insects typically harboring larger and more diversified P450 repertoires than specialists [36-39]. At the genomic level, these adaptive responses are largely mediated by gene duplication and gene birth-death dynamics, which provide the raw material for the expansion and diversification of insect P450 families [40, 41]. In addition, recent anthropogenic pressures such as extensive insecticide application may further shape the evolution and selection of detoxification-related genes in some insect lineages. For example, comparative genomics of the CYP9A subfamily in the noctuid pests Spodoptera frugiperda and S. exigua revealed species-specific gene duplications and sequence divergence within large CYP9A clusters [42]. Another well characterized case is the evolution of fenvalerate resistance in Australian Helicoverpa armigera, where a chimeric P450, CYP337B3 arose through gene duplication and recombination. This case illustrates how P450 gene birth-death dynamics can drive rapid adaptive evolution [43]. Additionally, P450 genes are frequently organized in tandem clusters, or near transposable element rich regions, which promote local duplication and rapid turnover through unequal crossing-over and recombination [23, 44, 45]. Moreover, regulatory evolution, including changes in inducibility, tissue specificity, and cis- or trans- regulatory signaling, provides an additional mechanism for rapid adaptation without the need for new genes [25]. Compared to vertebrates, insect P450 exhibits rapid evolution by high gene birth-death rates, sequence divergence, and lineage-specific innovation, particularly in environmental sensing, detoxification, and ecological interactions [40]. Across insect species, no clear correlation is observed between CYPome size and genome size (Fig. S1). For example, species with relatively large genomes (e.g., Locusta migratoria 6.5 Gb; Periplaneta americana 3.38 Gb; Diabrotica virgifera virgifera 2.42 Gb) exhibit substantial variation in CYPome size: while L. migratoria (94) and D. virgifera virgifera (105) have average-sized CYPomes, P. americana has one of the largest (178). Similarly, species with relatively small genomes (e.g., Culex quinquefasciatus 540 Mb, Tribolium castaneum 204 Mb) have some of the largest CYPomes (Fig. S1, Table S1). This pattern is consistent with previous studies showing that arthropod CYPomes are shaped by lineage-specific expansions (“bloom”) and dynamic birth-death processes, rather than uniform genome-wide expansion [19].
Classification and functional diversity of insect P450s
Despite extreme sequence diversity, insect CYP genes cluster into four conserved evolutionary clans: CYP3, CYP4, CYP2 and the mitochondrial clan. More recently, additional minor clans (e.g., clans 16 and 20) have been recognized in some lineages, although their functional roles remain largely unknown compared with the major CYP clans [19]. Table S1 summarizes CYP gene repertoires from 50 insect species representing major taxonomic groups, including agricultural pests, urban pests, and pollinators. Data were compiled from genomic studies with eight species represented by transcriptome-based datasets. The average insect CYPome size is 94 genes (Fig. 1; Table S1). Differences in CYPome size among insect orders are largely attributable to evolutionary dynamics and functional diversification of the four major CYP clans. The insect CYP3 clan represents the most expanded and diversified clan in insects [14, 40, 46]. The CYP3 clan comprises families such as CYP6, CYP9, CYP28, and many CYP300-400 series members and is most closely related evolutionarily to vertebrate CYP3 and CYP5 families. Members of this clan are frequently involved in xenobiotic metabolism, host-plant adaptation, and insecticide resistance, and are often organized in tandem gene clusters with tissue- and stage-specific expression patterns [14, 23, 40, 46]. Following duplication, many CYP3 paralogs may experience relaxed purifying selection, permitting sequence divergence and functional differentiation while retaining partial redundancy [47]. In T. castaneum, detoxification-related P450s, particularly within the CYP3 clans, have undergone extensive lineage-specific expansion. The T. castaneum genome encodes a markedly expanded repertoire of CYP3 members (27 subfamilies, 79 genes), especially within the CYP6 and CYP9 families [10].
Average cytochrome P450 (CYP) gene numbers across insect orders based on the species listed in Table S1, with bees and wasps shown separately. The y-axis shows CYPome size (number of CYP genes), and the x-axis shows insect orders. Bars represent the mean number of CYP genes per species within each group, categorized into the four major CYP clans (CYP2, mitochondrial, CYP3, and CYP4). The dashed line indicates the average CYPome size across all species analyzed.
Long-term exposure to diverse xenobiotics, particularly plant allelochemicals, has likely driven the retention and expansion of CYP3 gene clusters, while more recent exposure to synthetic insecticides has further selected for enhanced metabolic capacity [47, 48]. Lepidoptera provides a well-characterized example in which extensive P450 clustering reflects repeated tandem duplication driven by chronic exposure to chemically diverse host plants. Detoxification-related P450s, especially those in the CYP3 clans, evolve under a birth-death model characterized by high duplication and loss rates and relaxed functional constraints, favoring the accumulation of clustered paralogs [7, 42]. These clusters are frequently located in transposable element-rich genomic regions, which could facilitate local duplication via unequal crossing-over [44, 45]. In addition, physical clustering may confer functional and evolutionary advantages by maintaining groups of detoxification genes with complementary activities, enhancing the ability to cope with diverse allelochemicals and insecticides [49]. Variation in CYPome size may also reflect ecological adaptation. Polyphagous insects exposed to diverse host plant allelochemicals and environmental xenobiotics often exhibit expanded detoxification-related CYP families, whereas specialist species may retain more restricted CYP repertoires associated with narrower feeding niches. Mosquitoes (e.g., C. quinquefasciatus, Aedes aegypti, Anopheles gambiae) maintain relatively large CYP repertoires, likely reflecting the functional diversity of these enzymes in metabolizing insecticides, environmental pollutants, and host-derived compounds. Such diversity might confer metabolic flexibility and facilitate the rapid evolution of resistance under strong vector-control selection pressure [50]. Similarly, the American cockroach (P. americana) possesses one of the largest CYPomes reported in insects, primarily due to extensive expansion of the CYP3 clan genes. This expansion may enhance detoxification capacity towards diverse xenobiotics and support adaptation to chemically complex urban and waste-associated habitats, consistent with its highly generalist feeding ecology [51]. In contrast, extreme ecological specialization is correlated with CYPome contraction. Pediculus humanus (the human body louse) possesses one of the smallest CYP repertoires among insects, with only 12 CYP3 genes, consistent with long-term genome streamlining. As an obligate ectoparasite feeding exclusively on human blood, it encounters relatively limited chemical diversity and reduced exposure to environmental xenobiotics [52].
The CYP4 clan comprises the CYP4 family, which contains members from vertebrates and insects, as well as several families (e.g., CYP311-316) that can be traced back to ancestral genes in Caenorhabditis elegans [14]. CYP4 clan is highly diverse and typically represents the second-largest group in insect CYPomes [46]. For example, the T. castaneum genome encodes 47 individual genes belonging to 15 subfamilies in the CYP4 clan, with the CYP4 family being particularly expanded and comprising 27 genes (Fig. 1, Table S1) [10, 40]. However, the size of CYP4 clan varies remarkably among insect lineages, and substantial reductions are observed in some taxa. For example, most hymenopteran species harbor fewer than 10 CYP4 genes, while honey bee and other Apoidea only have 3-4 genes in CYP4 clan (Fig. 1, Table S1) [19]. Although many members remain poorly characterized, the CYP4 clan exhibits diverse functions in xenobiotic detoxification, cuticular hydrocarbon (CHC) biosynthesis, and fatty acid metabolism [14, 46]. Due to their central role in CHC biosynthesis, genes in CYP4G families are conserved across insect lineages with most species possessing at least one CYP4G gene [26, 53, 54]. For example, honey bee [55] and pea aphid [56] each carry a single CYP4G gene in their genomes, whereas Lepidopterans' genomes contain multiple CYP4G genes [57].
The mitochondrial CYP clan constitutes a distinct and highly conserved lineage within insect CYPomes. In contrast to plants, which lack mitochondrial P450s, insects and vertebrates retain this clan, although their functional repertoires differ significantly [40]. In vertebrates, mitochondrial P450s are primarily involved in steroid and vitamin D metabolism, whereas in insects they play central roles in ecdysteroid biosynthesis, hormone modification, and detoxification [14, 58]. As a monophyletic group localized to mitochondria, the mitochondria CYP clan includes highly conserved enzymes that are essential for ecdysteroid biosynthesis, including the Halloween genes CYP302, CYP314, and CYP315 [59]. While other mitochondrial P450s, particularly in dipterans (e.g. CYP12 family in house fly) are implicated in detoxification, resulting in highest number among other orders [40]. Another example is cat flea (Ctenocephalides felis), which has expanded CYP12 family in Mitochondria Clan, with 34 genes uncharacterized [60]. Recent study showed that some mitochondria P450s such as CYP333B3, CYP305B1, and CYP339A1 in H. armigera may also contribute to xenobiotic metabolism [58]. Overall, mitochondrial CYP clan size (average ~11 genes) is relatively stable across insect orders, with only modest expansion observed in Diptera and cat flea (Fig. 1, Table S1).
The insect CYP2 clan represents the most ancient P450 lineage and primarily functions in endogenous biosynthesis [46]. For example, CYP15, CYP18, and several CYP300-series families, are characterized by strong functional constraints and are implicated in hormone biosynthesis and development-related processes [40, 46]. However, recent studies have shown that some CYP2 clan P450s (e.g. CYP306 from Spodoptera litura, CYP305B, CYP18A1, and CYP303A1 from H. armigera) also participate in xenobiotic detoxification, in addition to their classical endogenous roles [58, 61]. The CYP2 clan contains an average of approximately 9 genes across insect orders (Fig. 1, Table S1) and exhibits relatively little inter-order variation compared with the highly dynamic CYP3 and CYP4 clans.
Structural diversity of insect P450s
Despite the central role of P450s in insect chemical adaptation, high-resolution three-dimensional structures of insect P450s remain scarce [2]. To date, most structural insights into P450 enzymes have been derived primarily from crystallographic and cryo-electron microscopy studies of bacterial, plant, and mammalian P450s, particularly human P450s [14, 62, 63]. Consequently, structural features of insect P450s are inferred mainly from comparative sequence analysis, homology modeling, and functional studies based on non-insect P450 structures. Nevertheless, the P450 superfamily is evolutionarily conserved, with core catalytic architectures shared across taxa, providing a solid framework for understanding the structure-function relationships of insect P450s involved in chemical adaptation [14, 62, 64].
In general, insect P450s exhibit a conserved global fold, comprising an N-terminal transmembrane domain linked to a larger catalytic domain that contains the heme-binding site responsible for catalysis (Fig. 2A) [65-68]. The catalytic domain adopts the classic P450 fold, characterized by a predominantly α-helical (~12 to 16 helices) with three to four β-sheets, and is organized into two sub-domains: the β domain and the α domain [65-70]. The secondary structural elements of the catalytic domain form a structural scaffold that supports both the active site and centrally located heme cofactor. In P450 structural studies, the hydrophobic transmembrane domain poses challenges for protein crystallization; consequently, most resolved structures lack the N-terminal transmembrane helix (TM helix) and focus on the soluble catalytic domain [71]. A notable exception is the full-length CYP51 class P450 from Saccharomyces cerevisiae solved by Monke et al. [71]. The S. cerevisiae CYP51A1 full-length structure provided evidence that the TM helix anchors P450 at the membrane without disrupting the canonical fold of the catalytic domain. The TM-helix spans the membrane, anchoring eukaryotic P450s to the endoplasmic reticulum. The catalytic domain faces the cytosol and interacts with substrates and redox partners, including cytochrome P450 reductase and cytochrome b5 [72-74]. P450 enzymes are membrane-anchored through the TM-helix, whereas the catalytic domain is only partially embedded in the lipid bilayer via hydrophobic regions such as the F/G loop and are largely exposed to the cytosol (Fig. 2A). This arrangement places the substrate access channel near the membrane interface, facilitating access to both cytosolic and lipophilic substrates [74, 75].
Structural model of an insect P450. The full-length amino acid sequence of CYP6BQ9 from the red flour beetle, Tribolium castaneum was used to generate an Alphafold 3 model [199]. CYP6BQ9 orientation within the endoplasmic reticulum (ER) was predicted using the PPM 3.0 server [200]. (A) Structural model of CYP6BQ9 shown in ribbon representation, with α-helices colored blue, β-strands gold, and loops grey. The N-terminal transmembrane (TM) helix is embedded in the lipid bilayer, and the catalytic domain extends into the cytosol. The heme cofactor is shown with carbon atoms colored tan and heteroatoms colored according to the CPK element scheme. Black horizontal lines indicate the membrane boundaries. Secondary structural elements are labeled. (B) Catalytic domain of the CYP6BQ9 model, colored and labeled as described in (A). Loops are rendered transparent to provide a clearer view of the heme center.
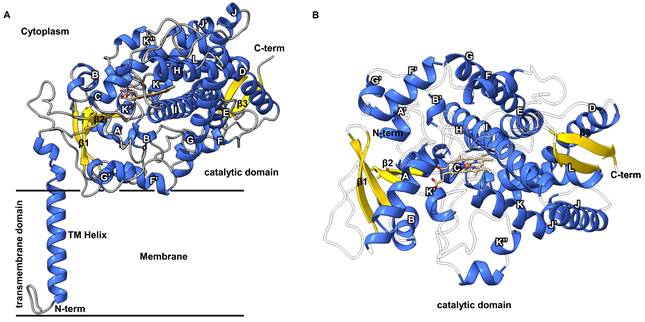
Insect P450s, including members of the CYP6, CYP9, and CYP4 families, are predicted to conform to the canonical P450 structure (Fig. 2A and B) [14, 76]. The structural core is formed by the conserved four helix bundle of D, E, I, and L. Leading into helix L is the heme-binding loop with the conserved PFxxGxRxCxG/A motif and the conserved cysteine that acts as the fifth ligand to the heme cofactor's iron, and a defining feature of all P450s [65, 69, 77]. This motif is conserved across insect P450 families, including detoxification-associated CYP6 and CYP9 enzymes, suggesting that their catalytic mechanisms resemble those of other eukaryotic and prokaryotic systems. There are five conserved P450 motifs that play central roles in structural fold stabilization, heme binding and/or catalysis. Besides PFxxGxRxCxG/A, the other motifs are WxxxR, GxE/DTT/S, ExxR, and PxxFxPE/DRF [14, 64, 78]. The WxxR motif is located on helix C, where the R residue binds one of the heme propionate groups. The GxE/DTT/S motif, located in the middle of helix I adjacent to the heme, contains a catalytically conserved threonine [64]. The ExxR motif, located in helix K, establishes a salt bridge network that stabilizes the overall protein structure [78]. Lastly, the proline-enriched PxxFxPE/DRF motif, located between the TM-helix and helix A, is thought to contribute to membrane positioning [78].
In contrast to the highly conserved heme-thiolate core, insect P450s exhibit substantial structural diversity within substrate recognition sites (SRSs), which are key determinants of substrate specificity and catalytic versatility. The six canonical SRS regions (SRS1-SRS6), primarily located in flexible structural elements such as the B′ helix, F-G region, and surrounding loops, display significant sequence and conformational variability among insect P450s [14, 79]. This structural plasticity enables the formation of diverse substrate-binding cavities and access channels, allowing insect P450s to accommodate a wide range of chemically distinct compounds, including plant allelochemicals and synthetic insecticides. Such diversity in SRS architecture is thought to underlie the remarkable adaptive capacity of insect P450s in response to environmental chemical challenges [80]. Comparative analyses indicate that SRS regions are among the most variable regions in insect P450s, particularly within expanded detoxification-related families such as CYP6 and CYP9. For example, CYP6 enzymes involved in pyrethroid or neonicotinoid metabolism often display marked sequence divergence in predicted SRS regions, consistent with differences in substrate specificity and catalytic efficiency [81]. The plasticity of the active-site cavity is a defining feature of P450 enzymes and plays a crucial role in insect chemical adaptation. Structural studies of mammalian P450s have shown that even modest amino acid substitutions within SRSs can significantly alter active-site volume, shape, and physicochemical properties [82, 83]. Amino acid substitutions within the SRS regions may alter substrate accessibility to the binding pocket or the range and catalytic efficiency of substrate metabolism, ultimately modulating the metabolic capacity of cytochrome P450 enzymes [80]. Although direct structural data for insect P450s remain limited, homology models of CYP6 and CYP9 enzymes support the hypothesis that adaptive changes in these regions facilitate the metabolism of chemically diverse compounds, including plant allelochemicals and synthetic insecticides [80, 84]. In vitro domain-swapping experiments involving SRS1 and SRS6 between CYP6AE17 and CYP6AE18 resulted in reciprocal changes in esfenvalerate-metabolizing activity. In addition, reciprocal exchange of the SRS1 regions between CYP6AE11 and CYP6AE20 caused improper folding of CYP6AE11/20 chimera, whereas the CYP6AE20/11 chimera acquired moderate esfenvalerate-metabolizing activity. These findings demonstrate that sequence variation within SRSs can influence protein folding, active-site conformation, and catalytic properties, thus contributing to the functional divergence of P450 enzymes [85]. In several insect species, amino acid substitutions within the predicted substrate-binding regions of P450 proteins (particularly CYP6) have been associated with insecticide resistance, illustrating how active-site plasticity enables adaptive detoxification [86, 87].
Functional versatility of insect P450s in chemical adaptation
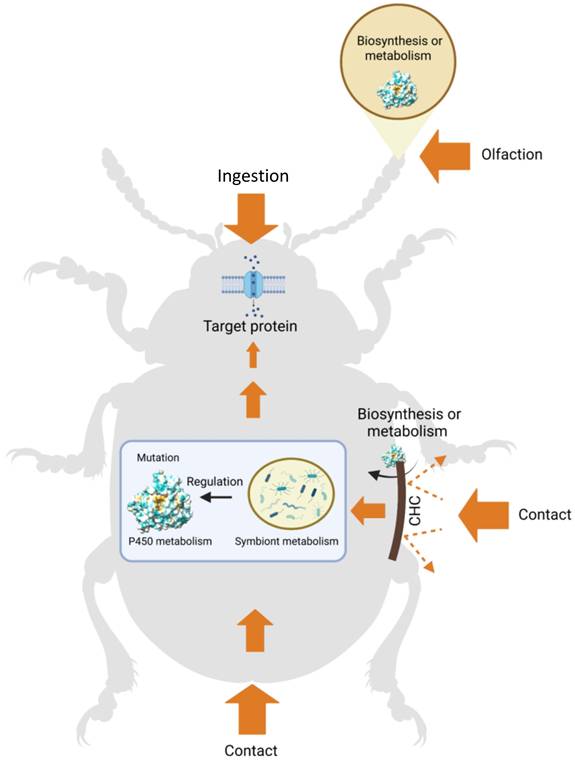
Cytochrome P450s are one of the most ancient and expansive enzyme superfamilies, known for catalyzing a wide range of oxidative transformations [23]. These enzymes mediate diverse reactions, including hydroxylation, epoxidation, and various dealkylation and oxidation processes involving oxygen, nitrogen, and sulfur atoms [14]. Their exceptional catalytic plasticity and broad substrate specificity enable insects to metabolize diverse endogenous and exogenous compounds [36]. As a result, P450s are key players in the detoxification and metabolic adaptation of insects to both naturally occurring toxins and synthetic xenobiotics, such as pesticides [22, 88]. In this review, we highlight the multifaceted roles of insect P450s in four major contexts of chemical adaptation: (1) P450-mediated host-plant adaptation and chemical communication, (2) P450-mediated detoxification of insecticides, (3) cuticle-associated resistance mechanisms, and (4) symbiont modulation of P450-mediated detoxification (Fig. 3).
Schematic overview of the diverse functions of insect cytochrome P450s and the evolutionary mechanisms underlying xenobiotic adaptation. Orange arrows indicate the direction and relative magnitude of chemical flux. Chemicals enter insects through multiple routes, including ingestion, antennal pores, cuticular penetration, and anal or ovipositional openings. The cuticle serves as a primary physical barrier to chemical entry, although cuticle-associated P450s may participate in the metabolism or biosynthesis of certain compounds. Chemicals entering through the antennae may also be metabolized by local P450s involved in olfactory processing. Compounds entering via ingestion, cuticular penetration, antennal exposure, or reproductive and anal routes may be metabolized by host P450s or symbiotic microorganisms before reaching their target sites. In addition, microbial symbionts may modulate host P450 expression and influence detoxification capacity. Figure created using BioRender.com.
Roles of P450s in host-plant adaptation and chemical communication
Insects and plants have interacted for hundreds of millions of years, influencing each other's evolution through long-term ecological and chemical relationships [9, 89]. As the primary food source for herbivorous insects, plants have evolved a diverse arsenal of allelochemicals to deter herbivory [36, 37]. These compounds include a wide range of insect repellents and toxins, such as alkaloids, terpenoids, phenolics, glucosinolates, cyanogenic glycosides, and proteinase inhibitors [12, 36, 90, 91]. In addition to direct-acting allelochemicals, plants employ indirect defenses through the release of herbivore-induced plant volatiles (HIPVs). Although chemically diverse, HIPVs primarily act as airborne signals that attract natural enemies of herbivores or prime defenses in nearby tissues and neighboring plants [92]. In response, herbivorous insects have developed diverse adaptive strategies to overcome plant chemical defenses, including behavioral avoidance, excretion, sequestration, enhanced metabolic detoxification, and target site insensitivity [9]. Among these mechanisms, metabolic detoxification by a variety of enzymes, such as P450s, glutathione S-transferases (GSTs), carboxylesterases (CCEs), and UDP-glycosyltransferases (UGTs), plays a central role in the neutralizing or metabolizing toxic compounds [22]. P450s are particularly critical among these enzymes due to their catalytic versatility, broad substrate specificity, and central role in initiating Phase I detoxification reactions [14, 46]. Plant allelochemicals are recognized as major evolutionary drivers of P450 diversification in phytophagous insects [9, 93-95]. In certain butterfly species, P450 duplications have enabled the detoxification of novel plant allelochemicals, facilitating host-plant shifts and potentially driving ecological divergence and speciation [95].
The evolutionary pressures driven by plant chemical defenses have also shaped different detoxification strategies among herbivorous insects with distinct feeding niches. Herbivorous insects are generally categorized as generalists, which feed on one or a few related plant taxa, or generalists, which consume a wide variety of plants [96]. This distinction often correlates with differences in detoxification capacity: specialists tend to evolve highly efficient P450s fine-tuned for a narrow set of allelochemicals, whereas generalists benefit from P450s with broader substrate specificities. For instance, the parsnip webworm (Depressaria pastinacella), a specialist feeding on furanocoumarin-rich Apiaceae plants, expresses the P450 enzyme CYP6AB3 [97]. Its variant, CYP6AB3v2, efficiently metabolizes imperatorin and myristicin, major host plant allelochemicals, highlighting the evolution of highly specialized detoxification mechanisms in specialist herbivores [98, 99]. In contrast, Helicoverpa zea, a highly polyphagous generalist, expresses CYP6B8, which is induced by allelochemicals such as xanthotoxin and exhibits broad substrate specificity, metabolizing diverse compounds including furanocoumarins and flavonoids [22, 100, 101]. Similarly, in the cotton bollworm, H. armigera, expansion of the CYP6AE subfamily enhances the capacity to metabolize diverse plant allelochemicals, contributing to its broad host range and dietary plasticity [94]. Functional divergence within P450 subfamilies may also contribute to species divergence. In a recent study, both the beet armyworm (S. exigua) and the fall armyworm (S. frugiperda) were shown to possess CYP9A subfamily members involved in allelochemical detoxification. CRISPR-mediated knockout of CYP9A genes in both species increases susceptibility to imperatorin and xanthotoxin, with species-specific differences in effect size and paralog involvement. These findings highlight lineage-specific functional divergence within the CYP9A subfamily that underlies adaptation to distinct host plant chemicals [42].
Importantly, P450-mediated adaptation to plant-derived toxins is not limited to herbivorous pests. Similar mechanisms are observed in pollinators such as the honey bee (Apis mellifera), which encounters a wide array of allelochemicals in its nectar, pollen, and propolis, and expresses multiple P450s (e.g. CYP6AS, CYP9Q, CYP336A1) capable of metabolizing flavonoids, alkaloids, and other plant secondary metabolites [102-104]. Notably, CYP4G11 and CYP9Q1-3 show task- and tissue-specific expression in honeybees. CYP4G11 is enriched in the antennae and prothoracic and mesothoracic legs of foragers, suggesting a role in chemosensory perception, whereas CYP9Q1-3 are most highly expressed in the metathoracic legs of foragers, supporting a role in detoxifying pollen phytochemicals [105].
In addition to their role in metabolizing plant toxins, insect P450s are increasingly recognized as odorant degrading enzymes in olfactory tissues, where they degrade volatile compounds and help reset chemoreception [106, 107]. Antennae are critical chemosensory organs responsible for detection and processing of environmental chemical cues. In this context, antennal P450s function to maintain chemosensory fidelity by rapidly degrading odorant molecules, thus preventing receptor overstimulation and ensuring signal clarity [107]. A striking example is the mountain pine beetle (Dendroctonus ponderosae), which encounters high concentrations of host-derived terpenoids [106]. In this species, P450s display remarkable tissue-specific expression patterns in response to chemically complex environments [108]. Additionally, cytochrome P450 reductase, the essential electron donor for all P450 genes, is also highly expressed in antennae of Colorado potato beetle and the common bed bug, suggesting that P450-mediated odorant degradation plays a significant role in olfactory tissues [109, 110]. P450s expressed in the gut, fat body, and/or Malpighian tubules primarily mediate xenobiotic detoxification, whereas antennal P450s may play multifunctional roles in olfaction, detoxification, and chemical communication [106, 107, 111]. For instance, the antenna-specific P450, CYP345E2, in the mountain pine beetle, D. ponderosae, metabolizes multiple monoterpenes, including (+)-3-carene, (±)-β-pinene, and (±)-α-pinene, suggesting its dual roles in odorant degradation and protection against the toxic effects of these volatiles [112, 113]. Interestingly, antennal P450s may also contribute to pheromone biosynthesis or degradation, highlighting their roles in chemical communication [108]. In the mountain pine beetles, CYP6DE1 is highly expressed in the antennae, heads, and fat bodies and in vitro assays show that CYP6DE1 catalyzes the conversion of α-pinene into trans-verbenol, an aggregation pheromone [114]. Similarly, in the red imported fire ant (Solenopsis invicta), CYP6K1 and CYP4V2 are highly expressed in adult worker antennae and are required for the detection of the alarm pheromone 2-ethyl-3,6(5)-dimethylpyrazine (EDMP). Silencing these genes via RNA interference reduces antennal electrophysiological responses and alters behavioral responses to EDMP, indicating that these P450s play critical roles in pheromone perception and signal processing [115]. Together, these examples illustrate the multifunctional roles of antennal P450s in insect olfaction, host-plant adaptation, and pheromone-mediated communication.
The evolutionary history of P450-mediated detoxification of plant chemicals may provide important insight into the origins and mechanisms of insecticide resistance. It has been proposed that some insects recruit enzymatic detoxification systems originally evolved for processing plant allelochemicals to metabolize synthetic insecticides [12, 116, 117]. This phenomenon, often referred to as 'pre-adaptation', provides a useful framework for understanding cross-resistance [9]. Indeed, the long evolutionary history of coping with plant secondary metabolites is thought to contribute to the rapid development of insecticide resistance in some species, mediated in part by shared detoxification pathways involving cytochrome P450s, GSTs, UGTs, and CCEs [118-121]. This hypothesis is further supported by the chemical similarities between certain plant-derived compounds and synthetic insecticides, many of which were developed based on natural plant toxins (e.g., pyrethrins and neonicotinoids) [9, 12, 122]. In addition, transcriptomic studies have revealed substantial overlap in gene expression responses to plant allelochemicals and synthetic insecticides, particularly in the up regulation of cytochrome P450s and other detoxification enzymes [120, 121, 123]. This convergence in detoxification machinery highlights an evolutionary continuum from adaptation to host plant chemistry to contemporary resistance toward synthetic pesticides.
Roles of P450s contribute to insecticide resistance and metabolism
P450s are among the most important enzymatic systems involved in the metabolism and detoxification of synthetic insecticides and play a central role in the development of insecticide resistance in disease vectors, agricultural and forest pests [3, 124, 125]. In addition, P450-mediated detoxification pathways contribute to chemical adaptation in non-target organisms, including both social and solitary pollinators [2, 14].
In mosquitoes, cytochrome P450s are major contributors to resistance against pyrethroids, which have been widely used in indoor residual spraying and long-lasting insecticidal nets [125]. Multiple resistance mechanisms have been identified in mosquitoes, with P450-mediated metabolic detoxification being among the most important [50, 126-129]. Overexpression of P450s involved in xenobiotic metabolism can arise through several mechanisms, including gene amplification at the genomic level and transcriptional upregulation in resistant strains [2, 128]. At the genomic level, copy number variance (CNV) is a common mechanism that increases the number of P450 gene copies, thereby enhancing detoxification capacity. For instance, CNVs in the Cyp6aa1-Cyp6p2 gene cluster have been documented in Anopheles species across Africa, conferring differential levels of resistance to pyrethroids and DDT compared with susceptible strains [130]. Beyond gene amplification, overexpression of P450 genes at the mRNA level can also be driven by cis- and/or trans- acting regulatory changes, including mutations in promoter regions and alterations in transcription factors [2, 131, 132]. Several key signaling pathways regulate insect P450 gene expression, including Cap'n'collar C/Nuclear factor erythroid 2-related factor 2-small Maf proteins (CncC/Nrf2-Maf) pathway, the G protein-coupled receptor (GPCR) pathway, and mitogen-activated protein kinase- cAMP responsive element-binding protein (MAPK-CREB) pathway [25]. In An. funestus, a 3-bp (AAC) deletion in the promoter region of CYP6P9b, located 50 bp upstream of a CncC/Nrf2-Maf binding site, has been associated with increased gene expression and pyrethroid resistance in southern African populations [128, 133, 134]. In C. quinquefasciatus, the GPCR signaling pathway has been implicated in P450-mediated permethrin resistance. Knockdown of four GPCR-related genes resulted in reduced expression of CYP9M10, CYP9J34, CYP6AA7, and CYP9J40 in a permethrin resistant strain, demonstrating the role of GPCR signaling in the regulation of P450 genes [135]. In addition to regulatory changes, recent studies have shown that point mutations in P450 coding sequences can directly alter enzyme activity and enhance catalytic efficiency. In An. funestus, site-directed mutagenesis revealed that three amino acid substitutions in CYP6P9b (Val109Ile, Asp335Glu, and Asn384Ser) are key resistance mutations responsible for enhanced pyrethroid metabolism in resistant alleles [136]. Similarly, a G454A mutation in CYP9K1 has been shown to increase metabolic activity toward the type II pyrethroid deltamethrin and contribute to elevated resistance in An. funestus populations [136, 137]. Together, these studies highlight how single or multiple amino acid substitutions in P450 enzymes can significantly alter catalytic efficiency and drive the evolution of insecticide resistance.
P450-mediated detoxification also plays a central role in the development of insecticide resistance in agricultural and forest pests. In the diamondback moth, Plutella xylostella (L.), a globally distributed pest of cruciferous crops, functional studies have shown that CYP6BG1 overexpression may contribute to chlorantraniliprole resistance [138]. Similarly, the Colorado potato beetle, Leptinotarsa decemlineata, relies heavily on P450-mediated detoxification as a key mechanism of imidacloprid resistance. Numerous P450s in the CYP4, CYP6, and CYP9 families are upregulated in resistant strains and can be induced by both host plant allelochemicals and insecticides [121]. The CncC/Nrf2-Maf signaling pathway has been identified controlling resistance-associated P450 expression in L. decemlineata and T. castaneum [139-141]. Additionally, microRNA (miRNA)-mediated regulation of P450 detoxification has been identified recently in whiteflies (Bemisia tabaci) and planthoppers (Nilaparvata lugens), providing new insights into the regulatory mechanisms underlying xenobiotic detoxification [142, 143]. In the forest pest Lymantria dispar, the methuselah-like GPCR gene (LdMthl1) and ocular albinism type 1 gene (LdOA1) regulate downstream CYP6 P450 genes, which are involved in deltamethrin resistance [144, 145]. Beyond gene amplification and regulatory mutations, emerging evidence suggests that epitranscriptomic regulation may also influence P450 expression and contribute to insecticide resistance. Chemical modifications of mRNA, such as N6-methyladenosine (m⁶A), have been implicated in modulating mRNA stability and translation efficiency, thereby potentially affecting detoxification capacity in insects. Two recent studies showed that m⁶A modification regulates the expression of CYP4C64 and CYP417B1, conferring resistance to thiamethoxam and imidacloprid, respectively [146, 147]. In addition, point mutations within the coding regions of P450 enzymes can also contribute to insecticide resistance by altering catalytic efficiency or substrate affinity. For example, an F116I mutation in CYP9A25 of Spodoptera litura has been identified as a key determinant of emamectin benzoate metabolism. This mutation, located within substrate recognition site 1 (SRS1), likely enhances metabolic activity by reducing steric hindrance and facilitating substrate access to the enzyme's active site [148]. Similarly, amino acid substitutions within the SRS regions of CYP6ER1 in the brown planthopper (N. lugens) have been shown to confer resistance to imidacloprid [87].
While cytochrome P450s often contribute to insecticide resistance in pests, they also play crucial roles in detoxifying agrochemicals in pollinators, helping to maintain pollinator health in pesticide-exposed environments [2, 149, 150]. Honey bees, once thought to be uniquely sensitive to pesticides due to relatively small repertoire of P450 genes [151], are now recognized as not necessarily among the most sensitive insect species [152, 153]. In fact, honey bees are generally more tolerant to agrichemicals, including neonicotinoids and fungicides, than many solitary bee species [154-156]. Several CYP9 enzymes have been functionally characterized for their detoxification roles in bees [2]. Notably, the CYP9Q family, including CYP9Q1-3 in honey bees and CYP9Q4/5 in bumblebees, effectively metabolizes the neonicotinoid thiacloprid [153]. Recent evidence suggests that CYP9Q2 has also shown to metabolize coumaphos, an organophosphate used to manage varroa mites in hives [157]. Moreover, CYP9Q2 and CYP9Q3 are involved in the detoxification of insecticides across a diverse range of chemical class, including diamide insecticide chlorantraniliprole and triazole fungicides [158]. However, P450-mediated insecticide detoxification capacity varies across bee lineages. In Megachilidae bees, species within the tribes Osmiini and Dioxyini possess CYP9BU genes capable of metabolizing thiacloprid [159], whereas the alfalfa leafcutter bee (Megachile rotundata) lacks CYP9Q-related P450s and substantially more sensitive to certain neonicotinoids [159, 160]. Detoxification in bees is a complex process involving not only P450s but also other enzymes, including GSTs, CCEs, UGTs, and ATP-binding cassette (ABC) transporters, as well as the microbiome, all of which may contribute to their adaptation to chemically intensive environments [150, 161]. Substantial future research is needed to fully understand the spectrum of genetic and symbiotic factors involved in this process.
Cuticle-associated P450s in chemical adaptation
The insect integument, composed sequentially from the innermost to outmost layer of the epidermis, endocuticle, exocuticle, and epicuticle, serves as the primary interface between the insect and its external environment. The outermost cuticular layer is coated with cuticular hydrocarbons (CHC), which plays a crucial role in protecting terrestrial insects from desiccation, and acting as signaling molecules in mating and communication [21, 162-165]. Accumulating evidence also indicates that CHCs contribute to insecticide resistance by reducing insecticide penetration [4, 26]. CYP4G enzymes catalyze the final step of CHC biosynthesis, converting long-chain fatty acyl-CoAs into hydrocarbons via alcohol and aldehyde intermediates [53]. In the lower termite Cryptotermes secundus, CHCs serve as queen pheromones. RNA interference (RNAi) silencing the CYP4C1 in queens significantly altered the royal CHC profile, leading to the loss of queen-specific scent and abolished queen recognition by workers [166]. While CYP4C1 is distinct from CYP4G enzymes, this study highlights the broader importance of cuticle-associated P450s in shaping CHC profiles that mediate chemical communication, a role that is mechanistically consistent with CYP4G-dependent hydrocarbon biosynthesis [26]. Additionally, Cyp301a1, a conserved P450 gene belonging to mitochondria clan, is involved in cuticle formation in insects. Although the signaling pathway is not clear, disrupting Cyp301a1 in D. melanogaster results in malformed abdominal cuticles [167].
Given the central role of CYP4G enzymes in CHC production and cuticle integrity, researchers have increasingly investigated their potential contributions to insecticide resistance. The first associations between CYP4G genes and insecticide resistance were reported in Blattella germanica insecticide-resistant strains [168]. This initial correlative evidence was subsequently supported by functional studies confirming a direct role for CYP4G genes in mediating resistance. In pyrethroid-resistant An. gambiae, for instance, overexpression of CYP4G16 contributes to thicker cuticular hydrocarbon layer compared with susceptible strain. Functional analyses demonstrated that CYP4G16, which is highly expressed in oenocytes, the primary cells responsible for CHC production, catalyzes the conversion of long-chain aldehydes into hydrocarbons, confirming its role as a hydrocarbon-forming decarbonylase [169]. Similarly, in pyrethroid-resistant B. germanica, elevated expression of CYP4G19 is associated with a thicker CHC layer and reduced cuticular permeability compared with susceptible strain. Conversely, RNAi -mediated knockdown of CYP4G19 increases the cuticle permeability and insecticide-induced mortality [170]. In N. lugens, RNAi-mediated silence of CYP4G76 and CYP4G115 decreases cuticular hydrocarbon thickness and desiccation tolerance in nymphs while enhancing penetration of pymetrozine, imidacloprid, thiamethoxam, and buprofezin [171].
Although most studies attribute CYP4G-associtated resistance to reduced insecticide penetration through enhanced CHC production, emerging evidence suggests that some cuticle-associated P450s may also contribute to insecticide resistance through direct metabolic detoxification. Evidence from Liriomyza trifolii suggests a potential direct metabolic role for CYP4G enzymes. Expression of CYP4G1 in Escherichia coli increased bacterial tolerance to abamectin in survival assays, although further biochemical characterization is needed to confirm direct xenobiotic metabolism by CYP4G enzymes [172]. Additional support for this possibility comes from studies in the bed bug, Cimex lectularius, where many detoxification genes, including several P450 genes highly expressed in pyrethroid-resistant strains, are predominantly expressed in the cuticle. This observation suggests that cuticle-associated detoxification may contribute to resistance [162].
Symbiont-mediated modulation of P450 detoxification
Microorganisms play a critical role in enabling insects to adapt to diverse environments, including natural, agricultural, and urban environments, reflecting highly dynamic host microbiome interactions [173-175]. To survive in chemically complex environments, insects rely on gut associated microbiomes contributing to digestion of polysaccharide, nutrient recycling, and xenobiotic detoxification [174, 175]. Although detoxification associated symbionts may not be essential for host growth or reproduction, they are crucial for facilitating chemical adaptation in insects [176].
One of the most direct ways in which symbionts contribute to chemical adaptation is through their ability to metabolize xenobiotics. Numerous studies have demonstrated that gut microbiomes can directly metabolize both plant toxins and synthetic pesticides [1, 177, 178]. For example, Pantoea spp., harbored by the cabbage stem flea beetle (Psylliodes chrysocephala), can degrade plant-derived toxic isothiocyanates in vitro. When beetles were treated with antibiotic, the isothiocyanate-degrading capacity was lost but recovered upon reintroduction of Pantoea, indicating that these symbionts contribute to host plant toxin detoxification [179]. Similarly, in Japanese sugarcane fields with long-term fenitrothion application, a fenitrothion-degrading microbiome, Burkholderia, was identified. Bean bug (Riptortus pedestris) populations harboring this symbiont showed significantly higher survival than those carrying non-degrading strains, demonstrating direct evidence that Burkholderia can confer pesticide resistance [176, 180]. Another example occurs in Grapholita molesta, where antibiotics-induced dysbiosis of the gut microbiota significantly increased larval sensitivity to emamectin benzoate. In addition, microbiota composition shifted when larvae moved from shoots to fruits, suggesting that dynamic microbiota communities modulate insecticide tolerance in response to dietary changes [181]. Together, these studies demonstrate that microbial symbionts can provide an immediate, metabolism-based mechanism for xenobiotic tolerance.
Beyond directly degrading xenobiotics, symbionts can also modulate host detoxification capacity by regulating endogenous detoxification systems, particularly cytochrome P450 enzymes [175, 177]. In A. mellifera, the gut microbiota promotes the expression of midgut P450s expression, and microbiome-deficient workers show significantly increased susceptibility to thiacloprid and tau-fluvalinate [182]. In mosquitoes, antibiotic treatment of Ae. aegypti larvae reduced P450 activity. In contrast, artificially increasing the abundance of the gut symbiont Serratia oryzae in Aedes albopictus larvae elevates P450 and other detoxification enzyme activities, thus improving survival following deltamethrin exposure [183, 184]. Similar patterns have been observed in planthoppers (N. lugens), where antibiotic treatment downregulated detoxification genes, including CYP4CE1 and CYP6ER1, and increased susceptibility to imidacloprid, chlorpyrifos, and clothianidin [185, 186]. However, symbionts do not universally enhance resistance. The symbiont Arsenophonus (S-type) reduced expression of CYP6AY1 and negatively affected imidacloprid resistance compared with the R-type strain, indicating that symbionts can also suppress host detoxification capacity. Such indirect modulation of host enzymatic system is associated with the downregulation of xenobiotic metabolism pathways [187].
In addition to direct and indirect detoxification mechanisms, hosts and their symbionts can cooperate to form integrated detoxification systems. Recent studies described a reciprocal host-symbiont detoxification strategy in which metabolic burdens are partitioned between partners [188]. In the bean bug (R. pedestris), the gut symbiont Burkholderia metabolized organophosphate insecticides into intermediate products. While the parent compound primarily affected the host insect, its metabolite, 3-methyl-4-nitrophenol (3M4N), was lethal to the symbiont. This conflict is resolved through rapid host-mediated excretion of the metabolite, thereby protecting the symbiont and maintaining detoxification capacity [188]. Such coordinated metabolic integration highlights how host physiological processes can complement symbiont-mediated xenobiotic metabolism, collectively enhancing pesticide tolerance.
Despite growing evidence for symbiont-mediated detoxification, the molecular mechanisms underlying how symbionts regulate host detoxification pathways in insects remain poorly understood. Nevertheless, insights from vertebrate systems may provide useful conceptual frameworks. RNA-seq comparisons between germ-free and conventionally reared mice have identified differential expression of cytochrome P450 enzymes and associated transcriptional regulators involved in drug metabolism [189, 190]. Microbial status and antibiotic exposure (i.e. metronidazole) can modulate transcription factors such as pregnane X receptor (PXR) and Nrf2, providing a mechanistic basis for differential cytochrome P450 expression [191, 192]. These findings suggest that microbial metabolites can modulate host signaling pathways that control cytochrome P450 expression. Future studies in insects should therefore focus on key transcriptional regulators and signaling pathways, including GPCRs, MAPK-CREB, AhR/ARNT, HR96, and CncC/Keap1, to elucidate how symbionts influence host detoxification processes [25].
While contributing to short-term physiological modulation, symbionts may also shape the long-term evolutionary trajectory of insect detoxification systems. Growing evidence suggests that microbial genes, including those encoding plant cell wall-degrading enzymes, have been horizontally transferred into insect genomes, representing an additional route for acquiring xenobiotic-degrading capabilities [193]. This challenges the traditional view that insect detoxification evolves exclusively through duplication and diversification of endogenous P450 genes [47]. Taken together, symbiont-mediated detoxification expands the functional and evolutionary landscape of insect P450-mediated chemical adaptation by integrating microbial metabolism, host regulatory plasticity, and genomic innovation.
Conclusion and future perspectives
Insect cytochrome P450s play central roles in chemical adaptation by facilitating insects to cope with diverse environmental challenges. Over the past 15 years, research on insect P450s has expanded rapidly, driven by advances in genome sequencing and annotation, alongside the maturation of functional genomics tools such as RNA interference (RNAi), CRISPR-based genome editing, and transgenic approaches [2]. These technologies have enabled systematic dissection of P450 function across diverse insect taxa and ecological contexts. Meanwhile, the development of heterologous expression systems has facilitated biochemical characterization of P450 enzymes and their roles in xenobiotic metabolism [2]. More recently, dedicated resources, including Insect-eP450DB [194], the Insect Cytochrome P450 Database [195], and the Arthropod P450 Enchiridion [196], have further accelerated discovery by integrating genomic, functional, and evolutionary data. These advances provide an increasingly comprehensive foundation for understanding how P450 diversification underpins insect chemical adaptation. Although insect genome sequencing has advanced significantly, many assemblies remain incomplete, particularly in repetitive regions such as centromeres and telomeres. As sequencing technologies improve toward near-complete genome assemblies, additional and potentially lineage-specific P450 genes and novel CYPomes, are likely to be discovered, which may further reveal functional diversification in detoxification pathways.
In future studies, artificial intelligence (AI) is expected to become an important tool in future P450 research [197]. Advances in AI-driven protein structure prediction and protein-ligand modeling may facilitate the identification of substrate recognition features, prediction of metabolic capabilities, and functional characterization of newly discovered P450 enzymes. In addition, AI-guided modeling may support the rational design of synergists targeting key detoxification P450s and contribute to the development of host-microbe co-metabolism frameworks for a more integrated understanding of xenobiotic detoxification. Future studies should also integrate structural biology, genome-editing technologies such as CRISPR/Cas, and applied pest management strategies to bridge molecular mechanisms with functional validation and practical applications [198]. Such multidisciplinary approaches will be essential for validating P450 function in vivo, elucidating the evolutionary dynamics of insecticide resistance, and translating mechanistic insights into actionable outcomes. These may include the development of novel synergists targeting key detoxification enzymes, as well as microbiome-informed and systems-based strategies for more sustainable pest and pollinator management.
Abbreviations
ABC transporters: ATP-binding cassette transporters; CHC: cuticular hydrocarbons; CncC: cap “n” collar isoform-C; CNV: copy number variance; COEs: carboxylesterases; EDMP: 2-ethyl-3,6(5)-dimethylpyrazine; GPCR: G protein-coupled receptor; GSTs: glutathione S-transferases; HIPVs: herbivore-induced plant volatiles; P450: Cytochrome P450 monooxygenase; RNAi: RNA interference; ROS: reactive oxygen species; SRS: substrate recognition site; UGTs: UDP-glycosyltransferases.
Supplementary Material
Supplementary figure and table.
Acknowledgements
This research was supported by a faculty start-up fund from Pennsylvania State University, NSF CAREER IOS-2144082, the USDA National Institute of Food and Federal Appropriations under Hatch Project #PEN04770 and Accession #1010058 (F.Z.). T.M. was supported by USDA NIFA postdoctoral fellowship, grant #2020-67034-31780/project accession#1022959 (2020-2022).
Author contributions
FZ developed the concept and framework of the manuscript. QC wrote the first draft. QC, TM, FZ summarized the table and made the figures. TM and FZ revised the manuscript. All authors approved the final manuscript.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Malook SU, Arora AK, Wong ACN. The role of microbiomes in shaping insecticide resistance: current insights and emerging paradigms. Curr Opin Insect Sci. 2025;69:101346
2. Nauen R, Bass C, Feyereisen R, Vontas J. The role of cytochrome P450s in insect toxicology and resistance. Annu Rev Entomol. 2022;67:105-24
3. Lu K, Song Y, Zeng R. The role of cytochrome P450-mediated detoxification in insect adaptation to xenobiotics. Curr Opin Insect Sci. 2021;43:103-7
4. Pu J, Chung H. New and emerging mechanisms of insecticide resistance. Curr Opin Insect Sci. 2024;63:101184
5. Gershenzon J, Dudareva N. The function of terpene natural products in the natural world. Nat Chem Biol. 2007;3:408-14
6. Dudareva N, Negre F, Nagegowda DA, Orlova I. Plant volatiles: recent advances and future perspectives. Crit Rev Plant Sci. 2006;25:417-40
7. Amezian D, Nauen R, Le Goff G. Comparative analysis of the detoxification gene inventory of four major Spodoptera pest species in response to xenobiotics. Insect Biochem Mol Biol. 2021;138:103646
8. Heidel-Fischer HM, Vogel H. Molecular mechanisms of insect adaptation to plant secondary compounds. Curr Opin Insect Sci. 2015;8:8-14
9. Després L, David JP, Gallet C. The evolutionary ecology of insect resistance to plant chemicals. Trends Ecol Evol. 2007;22:298-307
10. Zhu F, Moural TW, Shah K, Palli SR. Integrated analysis of cytochrome P450 gene superfamily in the red flour beetle, Tribolium castaneum. BMC Genomics. 2013;14:174
11. Liu N, Zhu F, Xu Q, Pridgeon JW, Gao X. Behavioral change, physiological modification, and metabolic detoxification: mechanisms of insecticide resistance. Acta Entomol Sin. 2006;49:671-9
12. Koirala BK S, Moural T, Zhu F. Functional and structural diversity of insect glutathione S-transferases in xenobiotic adaptation. Int J Biol Sci. 2022;18:5713-23
13. Cruse C, Moural TW, Zhu F. Dynamic roles of insect carboxyl/cholinesterases in chemical adaptation. Insects. 2023;14:194
14. Feyereisen R. Insect CYP genes and P450 enzymes. In: Gilbert LI, ed. Insect Molecular Biology and Biochemistry. San Diego: Academic Press. 2012:236-316
15. Liu N, Zhu F. House fly cytochrome P450s: their role in insecticide resistance and strategies in the isolation and characterization. In: Liu T, Kang L, editors. Recent Advances in Entomological Research: From Molecular Biology to Pest Management. New York: Springer-Verlag Berlin Heidelberg. 2011:246-57
16. Moural TW, Ban L, Hernandez JA, Wu M, Zhao C, Palli SR. et al. Silencing NADPH-cytochrome P450 reductase affects imidacloprid susceptibility, fecundity, and embryonic development in Leptinotarsa decemlineata. bioRxiv. 2020. 2020 09.29.318634
17. Shaik S, Cohen S, Wang Y, Chen H, Kumar D, Thiel W. P450 enzymes: their structure, reactivity, and selectivity-modeled by QM/MM calculations. Chem Rev. 2010;110:949-1017
18. Li F, Wang X, Zhou X. The genomics revolution drives a new era in entomology. Annu Rev Entomol. 2025;70:379-400
19. Dermauw W, Van Leeuwen T, Feyereisen R. Diversity and evolution of the P450 family in arthropods. Insect Biochem Mol Biol. 2020;127:103490
20. Nelson D. A world of cytochrome P450s. Philos Trans R Soc Lond B Biol Sci. 2013;368:20120430
21. Blomquist GJ, Ginzel MD. Chemical ecology, biochemistry, and molecular biology of insect hydrocarbons. Annu Rev Entomol. 2021;66:45-60
22. Li X, Schuler MA, Berenbaum MR. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu Rev Entomol. 2007;52:231-53
23. Feyereisen R. Insect P450 enzymes. Annu Rev Entomol. 1999;44:507-33
24. Katsavou E, Riga M, Ioannidis P, King R, Zimmer CT, Vontas J. Functionally characterized arthropod pest and pollinator cytochrome P450s associated with xenobiotic metabolism. Pestic Biochem Physiol. 2022;181:105005
25. Amezian D, Nauen R, Le Goff G. Transcriptional regulation of xenobiotic detoxification genes in insects -an overview. Pestic Biochem Physiol. 2021;174:104822
26. Feyereisen R. Origin and evolution of the CYP4G subfamily in insects, cytochrome P450 enzymes involved in cuticular hydrocarbon synthesis. Mol Phylogenet Evol. 2020;143:106695
27. Li X, Deng Z, Chen X. Regulation of insect P450s in response to phytochemicals. Curr Opin Insect Sci. 2021;43:108-16
28. Sezutsu H, Le Goff G, Feyereisen R. Origins of P450 diversity. Philos Trans R Soc Lond B Biol Sci. 2013;368:20120428
29. Lee DS, Nioche P, Hamberg M, Raman CS. Structural insights into the evolutionary paths of oxylipin biosynthetic enzymes. Nature. 2008;455:363-8
30. Nakahara K, Tanimoto T, Hatano K, Usuda K, Shoun H. Cytochrome P-450 55A1 (P-450dNIR) acts as nitric oxide reductase employing NADH as the direct electron donor. J Biol Chem. 1993;268:8350-5
31. Hrycay EG, Bandiera SM. Involvement of cytochrome P450 in reactive oxygen species formation and cancer. Adv Pharmacol. 2015;74:35-84
32. Lamb DC, Hargrove TY, Zhao B, Wawrzak Z, Goldstone JV, Nes WD. et al. Concerning P450 evolution: structural analyses support bacterial origin of sterol 14α-demethylases. Mol Biol Evol. 2020;38:952-67
33. Wollam J, Antebi A. Sterol regulation of metabolism, homeostasis, and development. Annu Rev Biochem. 2011;80:885-916
34. Scott GJ. Insect cytochrome P450s: thinking beyond detoxification. In: Liu N, ed. Recent Advances in Insect Physiology, Toxicology and Molecular Biology. Trivandrum, India: Research Signpost. 2008:117-124
35. Nelson DR. Metazoan cytochrome P450 evolution. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol. 1998;121:15-22
36. Schuler MA. The role of cytochrome P450 monooxygenases in plant-insect interactions. Plant Physiol. 1996;112:1411-9
37. Schuler MA. P450s in plant-insect interactions. Biochim Biophys Acta. 2011;1814:36-45
38. Berenbaum MR, Zangerl AR. Facing the future of plant-insect interaction research: le retour à la "raison d'être". Plant Physiol. 2008;146:804-11
39. Rane RV, Ghodke AB, Hoffmann AA, Edwards OR, Walsh TK, Oakeshott JG. Detoxifying enzyme complements and host use phenotypes in 160 insect species. Curr Opin Insect Sci. 2019;31:131-8
40. Feyereisen R. Arthropod CYPomes illustrate the tempo and mode in P450 evolution. Biochim Biophys Acta. 2011;1814:19-28
41. Calla B, Noble K, Johnson RM, Walden KKO, Schuler MA, Robertson HM. et al. Cytochrome P450 diversification and hostplant utilization patterns in specialist and generalist moths: Birth, death and adaptation. Mol Ecol. 2017;26:6021-35
42. Shi Y, Liu Q, Lu W, Yuan J, Yang Y, Oakeshott J. et al. Divergent amplifications of CYP9A cytochrome P450 genes provide two noctuid pests with differential protection against xenobiotics. Proc Natl Acad Sci U S A. 2023;120:e2308685120
43. Joußen N, Agnolet S, Lorenz S, Schöne SE, Ellinger R, Schneider B. et al. Resistance of Australian Helicoverpa armigera to fenvalerate is due to the chimeric P450 enzyme CYP337B3. Proc Natl Acad Sci U S A. 2012;109:15206-11
44. You M, Yue Z, He W, Yang X, Yang G, Xie M. et al. A heterozygous moth genome provides insights into herbivory and detoxification. Nat Genet. 2013;45:220-5
45. Chen S, Li X. Transposable elements are enriched within or in close proximity to xenobiotic-metabolizing cytochrome P450 genes. BMC Evol Biol. 2007;7:46
46. Feyereisen R. Evolution of insect P450. Biochem Soc Trans. 2006;34:1252-5
47. Zhang J. Evolution by gene duplication: an update. Trends Ecol Evol. 2003;18:292-8
48. Wondji CS, Irving H, Morgan J, Lobo NF, Collins FH, Hunt RH. et al. Two duplicated P450 genes are associated with pyrethroid resistance in Anopheles funestus, a major malaria vector. Genome Res. 2009;19:452-9
49. Wang H, Shi Y, Wang L, Liu S, Wu S, Yang Y. et al. CYP6AE gene cluster knockout in Helicoverpa armigera reveals role in detoxification of phytochemicals and insecticides. Nat Commun. 2018;9:4820
50. David JP, Ismail HM, Chandor-Proust A, Paine MJ. Role of cytochrome P450s in insecticide resistance: impact on the control of mosquito-borne diseases and use of insecticides on Earth. Philos Trans R Soc Lond B Biol Sci. 2013;368:20120429
51. Li S, Zhu S, Jia Q, Yuan D, Ren C, Li K. et al. The genomic and functional landscapes of developmental plasticity in the American cockroach. Nat Commun. 2018;9:1008
52. Kirkness EF, Haas BJ, Sun W, Braig HR, Perotti MA, Clark JM. et al. Genome sequences of the human body louse and its primary endosymbiont provide insights into the permanent parasitic lifestyle. Proc Natl Acad Sci U S A. 2010;107:12168-73
53. Qiu Y, Tittiger C, Wicker-Thomas C, Le Goff G, Young S, Wajnberg E. et al. An insect-specific P450 oxidative decarbonylase for cuticular hydrocarbon biosynthesis. Proc Natl Acad Sci U S A. 2012;109:14858-63
54. Holze H, Schrader L, Buellesbach J. Advances in deciphering the genetic basis of insect cuticular hydrocarbon biosynthesis and variation. Heredity. 2021;126:219-34
55. Calla B, MacLean M, Liao LH, Dhanjal I, Tittiger C, Blomquist GJ. et al. Functional characterization of CYP4G11-a highly conserved enzyme in the western honey bee Apis mellifera. Insect Mol Biol. 2018;27:661-74
56. Chen N, Fan YL, Bai Y, Li XD, Zhang ZF, Liu TX. Cytochrome P450 gene, CYP4G51, modulates hydrocarbon production in the pea aphid, Acyrthosiphon pisum. Insect Biochem Mol Biol. 2016;76:84-94
57. Xin Y, Chen N, Wang Y, Ni R, Zhao H, Yang P. et al. CYP4G8 is responsible for the synthesis of methyl-branched hydrocarbons in the polyphagous caterpillar of Helicoverpa armigera. Insect Biochem Mol Biol. 2022;140:103701
58. Shi Y, Qu Q, Wang C, He Y, Yang Y, Wu Y. Involvement of CYP2 and mitochondrial clan P450s of Helicoverpa armigera in xenobiotic metabolism. Insect Biochem Mol Biol. 2022;140:103696
59. Iga M, Kataoka H. Recent studies on insect hormone metabolic pathways mediated by cytochrome P450 enzymes. Biol Pharm Bull. 2012;35:838-43
60. Feyereisen R. The P450 genes of the cat flea. Curr Res Insect Sci. 2022;2:100032
61. Liu J, Hua J, Wang Y, Guo X, Luo S. Caterpillars detoxify diterpenoid from Nepeta stewartiana by the molting hormone gene CYP306A1. J Agric Food Chem. 2023;71:10670-82
62. Guengerich FP, Waterman MR, Egli M. Recent structural insights into cytochrome P450 function. Trends Pharmacol Sci. 2016;37:625-40
63. Su M, Chakraborty S, Osawa Y, Zhang H. Cryo-EM reveals the architecture of the dimeric cytochrome P450 CYP102A1 enzyme and conformational changes required for redox partner recognition. J Biol Chem. 2020;295:1637-45
64. Denisov IG, Makris TM, Sligar SG, Schlichting I. Structure and chemistry of cytochrome P450. Chem Rev. 2005;105:2253-78
65. Hasemann CA, Kurumbail RG, Boddupalli SS, Peterson JA, Deisenhofer J. Structure and function of cytochromes P450: a comparative analysis of three crystal structures. Structure. 1995;3:41-62
66. Ravichandran KG, Boddupalli SS, Hasermann CA, Peterson JA, Deisenhofer J. Crystal structure of hemoprotein domain of P450BM-3, a prototype for microsomal P450's. Science. 1993;261:731-6
67. Williams PA, Cosme J, Sridhar V, Johnson EF, McRee DE. Microsomal cytochrome P450 2C5: comparison to microbial P450s and unique features. J Inorg Biochem. 2000;81:183-90
68. Yano JK, Wester MR, Schoch GA, Griffin KJ, Stout CD, Johnson EF. The structure of human microsomal cytochrome P450 3A4 determined by X-ray crystallography to 2.05-Å Resolution. J Biol Chem. 2004;279:38091-4
69. Werck-Reichhart D, Feyereisen R. Cytochromes P450: a success story. Genome Biol. 2000;1:reviews3003
70. Zhang B, Lewis KM, Abril A, Davydov DR, Vermerris W, Sattler SE. et al. Structure and function of the cytochrome P450 monooxygenase cinnamate 4-hydroxylase from Sorghum bicolor. Plant Physiol. 2020;183:957-73
71. Monk BC, Tomasiak TM, Keniya MV, Huschmann FU, Tyndall JDA, O'Connell JD. et al. Architecture of a single membrane spanning cytochrome P450 suggests constraints that orient the catalytic domain relative to a bilayer. Proc Natl Acad Sci U S A. 2014;111:3865-70
72. Black SD, Martin ST, Smith CA. Membrane topology of liver microsomal cytochrome P450 2B4 determined via monoclonal antibodies directed to the halt-transfer signal. Biochemistry. 1994;33:6945-51
73. Cojocaru V, Balali-Mood K, Sansom MS, Wade RC. Structure and dynamics of the membrane-bound cytochrome P450 2C9. PLoS Comput Biol. 2011;7:e1002152
74. Mukherjee G, Nandekar PP, Wade RC. An electron transfer competent structural ensemble of membrane- bound cytochrome P450 1A1 and cytochrome P450 oxidoreductase. Commun Biol. 2021;4:55
75. Baylon JL, Lenov IL, Sligar SG, Tajkhorshid E. Characterizing the membrane-bound state of cytochrome P450 3A4: structure, depth of insertion, and orientation. J Am Chem Soc. 2013;135:8542-51
76. Lertkiatmongkol P, Jenwitheesuk E, Rongnoparut P. Homology modeling of mosquito cytochrome P450 enzymes involved in pyrethroid metabolism: insights into differences in substrate selectivity. BMC Res Notes. 2011;4:321
77. Lu J, Zhang H, Wang Q, Huang X. Genome-wide identification and expression pattern of cytochrome P450 genes in the social aphid Pseudoregma bambucicola. Insects. 2023;14:212
78. Fang Y, Tai Z, Hu K, Luo L, Yang S, Liu M. et al. Comprehensive review on plant cytochrome P450 evolution: copy number, diversity, and motif analysis from chlorophyta to dicotyledoneae. Genome Biol Evol. 2024;16:evae240
79. Di Nardo G, Gilardi G. Natural compounds as pharmaceuticals: the key role of cytochromes P450 reactivity. Trends Biochem Sci. 2020;45:511-25
80. Schuler MA, Berenbaum MR. Structure and function of cytochrome P450s in insect adaptation to natural and synthetic toxins: insights gained from molecular modeling. J Chem Ecol. 2013;39:1232-45
81. Xiao T, Lu K. Functional characterization of CYP6AE subfamily P450s associated with pyrethroid detoxification in Spodoptera litura. Int J Biol Macromol. 2022;219:452-62
82. Domanski TL, Halpert JR. Analysis of mammalian cytochrome P450 structure and function by site-directed mutagenesis. Curr Drug Metab. 2001;2:117-37
83. Williams PA, Cosme J, Vinkovic DM, Ward A, Angove HC, Day PJ. et al. Crystal structures of human cytochrome P450 3A4 bound to metyrapone and progesterone. Science. 2004;305:683-6
84. Guo Y, Zhang X, Wu H, Yu R, Zhang J, Zhu KY. et al. Identification and functional analysis of a cytochrome P450 gene CYP9AQ2 involved in deltamethrin detoxification from Locusta migratoria. Pestic Biochem Physiol. 2015;122:1-7
85. Shi Y, O'Reilly AO, Sun S, Qu Q, Yang Y, Wu Y. Roles of the variable P450 substrate recognition sites SRS1 and SRS6 in esfenvalerate metabolism by CYP6AE subfamily enzymes in Helicoverpa armigera. Insect Biochem Mol Biol. 2020;127:103486
86. Duarte A, Pym A, Garrood WT, Troczka BJ, Zimmer CT, Davies TGE. et al. P450 gene duplication and divergence led to the evolution of dual novel functions and insecticide cross-resistance in the brown planthopper Nilaparvata lugens. PLoS Genet. 2022;18:e1010279
87. Zimmer CT, Garrood WT, Singh KS, Randall E, Lueke B, Gutbrod O. et al. Neofunctionalization of duplicated P450 genes drives the evolution of insecticide resistance in the brown planthopper. Curr Biol. 2018;28:268-74
88. Scott JG, Liu N, Wen Z. Insect cytochromes P450: diversity, insecticide resistance and tolerance to plant toxins. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol. 1998;121:147-55
89. Fürstenberg-Hägg J, Zagrobelny M, Bak S. Plant defense against insect herbivores. Int J Mol Sci. 2013;14:10242-97
90. Fraenkel GS. The raison d'ĕtre of secondary plant substances; these odd chemicals arose as a means of protecting plants from insects and now guide insects to food. Science. 1959;129:1466-70
91. Howe GA, Jander G. Plant immunity to insect herbivores. Annu Rev Plant Biol. 2008;59:41-66
92. War AR, Taggar GK, Hussain B, Taggar MS, Nair RM, Sharma HC. Plant defence against herbivory and insect adaptations. AoB PLANTS. 2018 10
93. Alyokhin A, Chen YH. Adaptation to toxic hosts as a factor in the evolution of insecticide resistance. Curr Opin Insect Sci. 2017;21:33-8
94. Shi Y, Wang H, Liu Z, Wu S, Yang Y, Feyereisen R. et al. Phylogenetic and functional characterization of ten P450 genes from the CYP6AE subfamily of Helicoverpa armigera involved in xenobiotic metabolism. Insect Biochem Mol Biol. 2018;93:79-91
95. Edger PP, Heidel-Fischer HM, Bekaert M, Rota J, Glöckner G, Platts AE. et al. The butterfly plant arms-race escalated by gene and genome duplications. Proc Natl Acad Sci U S A. 2015;112:8362-6
96. Ali JG, Agrawal AA. Specialist versus generalist insect herbivores and plant defense. Trends Plant Sci. 2012;17:293-302
97. Mao W, Rupasinghe S, Zangerl AR, Schuler MA, Berenbaum MR. Remarkable substrate-specificity of CYP6AB3 in Depressaria pastinacella, a highly specialized caterpillar. Insect Mol Biol. 2006;15:169-79
98. Mao W, Rupasinghe SG, Zangerl AR, Berenbaum MR, Schuler MA. Allelic variation in the Depressaria pastinacella CYP6AB3 protein enhances metabolism of plant allelochemicals by altering a proximal surface residue and potential interactions with cytochrome P450 reductase. J Biol Chem. 2007;282:10544-52
99. Mao W, Zangerl AR, Berenbaum MR, Schuler MA. Metabolism of myristicin by Depressaria pastinacella CYP6AB3v2 and inhibition by its metabolite. Insect Biochem Mol Biol. 2008;38:645-51
100. Li X, Baudry J, Berenbaum MR, Schuler MA. Structural and functional divergence of insect CYP6B proteins: from specialist to generalist cytochrome P450. Proc Natl Acad Sci U S A. 2004;101:2939-44
101. Rupasinghe SG, Wen Z, Chiu TL, Schuler MA. Helicoverpa zea CYP6B8 and CYP321A1: different molecular solutions to the problem of metabolizing plant toxins and insecticides. Protein Eng Des Sel. 2007;20:615-24
102. Mao W, Rupasinghe SG, Johnson RM, Zangerl AR, Schuler MA, Berenbaum MR. Quercetin-metabolizing CYP6AS enzymes of the pollinator Apis mellifera (Hymenoptera: Apidae). Comp Biochem Physiol B Biochem Mol Biol. 2009;154:427-34
103. Mao W, Schuler MA, Berenbaum MR. Honey constituents up-regulate detoxification and immunity genes in the western honey bee Apis mellifera. Proc Natl Acad Sci U S A. 2013;110:8842-6
104. Haas J, Beck E, Troczka BJ, Hayward A, Hertlein G, Zaworra M. et al. A conserved hymenopteran-specific family of cytochrome P450s protects bee pollinators from toxic nectar alkaloids. Sci Adv. 2023;9:eadg0885
105. Mao W, Schuler MA, Berenbaum MR. Task-related differential expression of four cytochrome P450 genes in honeybee appendages. Insect Mol Biol. 2015;24:582-8
106. Seybold SJ, Huber DPW, Lee JC, Graves AD, Bohlmann J. Pine monoterpenes and pine bark beetles: a marriage of convenience for defense and chemical communication. Phytochem Rev. 2006;5:143-78
107. Leal WS. Odorant reception in insects: roles of receptors, binding proteins, and degrading enzymes. Annu Rev Entomol. 2013;58:373-91
108. Chiu CC, Keeling CI, Bohlmann J. Cytochromes P450 preferentially expressed in antennae of the mountain pine beetle. J Chem Ecol. 2019;45:178-86
109. Cruse C. Behavioral and functional characterization of xenobiotic responsible enzymes in the Colorado potato beetle, Leptinotarsa decemlineata. Pennsylvania State University. 2023
110. Zhu F, Sams S, Moural T, Haynes KF, Potter MF, Palli SR. RNA interference of NADPH-cytochrome P450 reductase results in reduced insecticide resistance in the bed bug, Cimex lectularius. PLoS One. 2012;7:e31037
111. Wu H, Liu Y, Shi X, Zhang X, Ye C, Zhu KY. et al. Transcriptome analysis of antennal cytochrome P450s and their transcriptional responses to plant and locust volatiles in Locusta migratoria. Int J Biol Macromol. 2020;149:741-53
112. Keeling CI, Henderson H, Li M, Dullat HK, Ohnishi T, Bohlmann J. CYP345E2, an antenna-specific cytochrome P450 from the mountain pine beetle, Dendroctonus ponderosae Hopkins, catalyses the oxidation of pine host monoterpene volatiles. Insect Biochem Mol Biol. 2013;43:1142-1151
113. Chiu CC, Keeling CI, Bohlmann J. Toxicity of pine monoterpenes to mountain pine beetle. Sci Rep. 2017;7:8858
114. Chiu CC, Keeling CI, Bohlmann J. The cytochrome P450 CYP6DE1 catalyzes the conversion of α-pinene into the mountain pine beetle aggregation pheromone trans-verbenol. Sci Rep. 2019;9:1477
115. Jiang X, Shen J, Lin P, Hou Y. High antennal expression of CYP6K1 and CYP4V2 participate in the recognition of alarm pheromones by Solenopsis invicta Buren. Insects. 2025 16
116. Gordon HT. Nutritional factors in insect resistance to chemicals. Annu Rev Entomol. 1961;6:27-54
117. Hardy NB, Peterson DA, Ross L, Rosenheim JA. Does a plant-eating insect's diet govern the evolution of insecticide resistance? comparative tests of the pre-adaptation hypothesis. Evol Appl. 2018;11:739-47
118. Moural TW, Hernandez JA, Chen Q-R, Koirala Bk S, Liu Y, Cofer TM. et al. Crystal structure and ligand binding of a sigma-class glutathione S-transferase associated with cross-resistance in a specialist herbivore. Int J Biol Macromol. 2025;323:147108
119. Morales MA, Mendoza BM, Lavine LC, Lavine MD, Walsh DB, Zhu F. Selection of reference genes for expression studies of xenobiotic adaptation in Tetranychus urticae. Int J Biol Sci. 2016;12:1129-39
120. Dermauw W, Wybouw N, Rombauts S, Menten B, Vontas J, Grbic M. et al. A link between host plant adaptation and pesticide resistance in the polyphagous spider mite Tetranychus urticae. Proc Natl Acad Sci U S A. 2013;110:E113-22
121. Zhu F, Moural TW, Nelson DR, Palli SR. A specialist herbivore pest adaptation to xenobiotics through up-regulation of multiple cytochrome P450s. Sci Rep. 2016;6:20421
122. Crossley MS, Snyder WE, Hardy NB. Insect-plant relationships predict the speed of insecticide adaptation. Evol Appl. 2020;14:290-6
123. Wen Z, Zeng RS, Niu G, Berenbaum MR, Schuler MA. Ecological significance of induction of broad-substrate cytochrome P450s by natural and synthetic inducers in Helicoverpa zea. J Chem Ecol. 2009;35:183-9
124. Adesanya AW, Lavine MD, Moural TW, Lavine LC, Zhu F, Walsh DB. Mechanisms and management of acaricide resistance for Tetranychus urticae in agroecosystems. J Pest Sci. 2021;94:639-63
125. Vontas J, Kioulos E, Pavlidi N, Morou E, della Torre A, Ranson H. Insecticide resistance in the major dengue vectors Aedes albopictus and Aedes aegypti. Pestic Biochem Physiol. 2012;104:126-31
126. Smith LB, Kasai S, Scott JG. Pyrethroid resistance in Aedes aegypti and Aedes albopictus: important mosquito vectors of human diseases. Pestic Biochem Physiol. 2016;133:1-12
127. Moyes CL, Vontas J, Martins AJ, Ng LC, Koou SY, Dusfour I. et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl Trop Dis. 2017;11:e0005625
128. Vontas J, Katsavou E, Mavridis K. Cytochrome P450-based metabolic insecticide resistance in Anopheles and Aedes mosquito vectors: muddying the waters. Pestic Biochem Physiol. 2020;170:104666
129. Ranson H, N'guessan R, Lines J, Moiroux N, Nkuni Z, Corbel V. Pyrethroid resistance in African anopheline mosquitoes: what are the implications for malaria control? Trends Parasitol. 2011;27:91-8
130. Lucas ER, Miles A, Harding NJ, Clarkson CS, Lawniczak MKN, Kwiatkowski DP. et al. Whole-genome sequencing reveals high complexity of copy number variation at insecticide resistance loci in malaria mosquitoes. Genome Res. 2019;29:1250-61
131. Liu N. Insecticide resistance in mosquitoes: impact, mechanisms, and research directions. Annu Rev Entomol. 2015;60:537-59
132. Liu N, Li M, Gong Y, Liu F, Li T. Cytochrome P450s - their expression, regulation, and role in insecticide resistance. Pestic Biochem Physiol. 2015;120:77-81
133. Mugenzi LMJ, Menze BD, Tchouakui M, Wondji MJ, Irving H, Tchoupo M. et al. Cis-regulatory CYP6P9b P450 variants associated with loss of insecticide-treated bed net efficacy against Anopheles funestus. Nat Commun. 2019;10:4652
134. Weedall GD, Mugenzi LMJ, Menze BD, Tchouakui M, Ibrahim SS, Amvongo-Adjia N. et al. A cytochrome P450 allele confers pyrethroid resistance on a major African malaria vector, reducing insecticide-treated bed net efficacy. Sci Transl Med. 2019 11
135. Li T, Liu L, Zhang L, Liu N. Role of G-protein-coupled receptor-related genes in insecticide resistance of the mosquito, Culex quinquefasciatus. Sci Rep. 2014;4:6474
136. Ibrahim SS, Riveron JM, Bibby J, Irving H, Yunta C, Paine MJ. et al. Allelic variation of cytochrome P450s drives resistance to bednet insecticides in a major malaria vector. PLoS Genet. 2015;11:e1005618
137. Djoko Tagne CS, Kouamo MFM, Tchouakui M, Muhammad A, Mugenzi LJL, Tatchou-Nebangwa NMT. et al. A single mutation G454A in the P450 CYP9K1 drives pyrethroid resistance in the major malaria vector Anopheles funestus reducing bed net efficacy. Genetics. 2025;229:1-40
138. Li X, Li R, Zhu B, Gao X, Liang P. Overexpression of cytochrome P450 CYP6BG1 may contribute to chlorantraniliprole resistance in Plutella xylostella (L.). Pest Manag Sci. 2018;74:1386-93
139. Zhu F, Parthasarathy R, Bai H, Woithe K, Kaussmann M, Nauen R. et al. A brain-specific cytochrome P450 responsible for the majority of deltamethrin resistance in the QTC279 strain of Tribolium castaneum. Proc Natl Acad Sci U S A. 2010;107:8557-62
140. Kalsi M, Palli SR. Transcription factors, CncC and Maf, regulate expression of CYP6BQ genes responsible for deltamethrin resistance in Tribolium castaneum. Insect Biochem Mol Biol. 2015;65:47-56
141. Kalsi M, Palli SR. Transcription factor cap n collar C regulates multiple cytochrome P450 genes conferring adaptation to potato plant allelochemicals and resistance to imidacloprid in Leptinotarsa decemlineata (Say). Insect Biochem Mol Biol. 2017;83:1-12
142. Zhao X, Chen X, He M, Wu S, Shi Y, Luo D. et al. miRNAs modulate altered expression of cytochrome P450s and nicotinic acetylcholine receptor subunits conferring both metabolic and target resistance to sulfoxaflor in Nilaparvata lugens (Stål). Int J Biol Macromol. 2025;290:138992
143. Gong PP, Wei XG, Liu SN, Yang J, Fu BL, Liang JJ. et al. Novel_miR-1517 mediates CYP6CM1 to regulate imidacloprid resistance in Bemisia tabaci (Hemiptera: Gennadius). Pestic Biochem Physiol. 2023;194:105469
144. Cao C, Sun L, Du H, Moural TW, Bai H, Liu P. et al. Physiological functions of a methuselah-like G protein coupled receptor in Lymantria dispar Linnaeus. Pestic Biochem Physiol. 2019;160:1-10
145. Sun L, Liu P, Zhang C, Du H, Wang Z, Moural TW. et al. Ocular albinism type 1 regulates deltamethrin tolerance in Lymantria dispar and Drosophila melanogaster. Front Physiol. 2019;10:766
146. Yang X, Wei X, Yang J, Du T, Yin C, Fu B. et al. Epitranscriptomic regulation of insecticide resistance. Sci Adv. 2021 7
147. Yang J, Xue H, Huang M, Wei X, Zhang R, Guo Z. et al. m6A mediated regulation of CYP417B1 contributes to imidacloprid resistance in Bemisia tabaci. J Agric Food Chem. 2025;73:16766-77
148. Shi Y, Su T, Yu Z, Mei W, Wu Y, Yang Y. F116I mutation in CYP9A25 associated with resistance to emamectin benzoate in Spodoptera litura. Pest Manag Sci. 2025;81:247-54
149. Berenbaum MR, Johnson RM. Xenobiotic detoxification pathways in honey bees. Curr Opin Insect Sci. 2015;10:51-8
150. Moural TW, Koirala B K S, Bhattarai G, He Z, Guo H, Phan NT. et al. Architecture and potential roles of a delta-class glutathione S-transferase in protecting honey bee from agrochemicals. Chemosphere. 2024;350:141089
151. Claudianos C, Ranson H, Johnson RM, Biswas S, Schuler MA, Berenbaum MR. et al. A deficit of detoxification enzymes: pesticide sensitivity and environmental response in the honeybee. Insect Mol Biol. 2006;15:615-36
152. Hardstone MC, Scott JG. Is Apis mellifera more sensitive to insecticides than other insects? Pest Manag Sci. 2010;66:1171-80
153. Manjon C, Troczka BJ, Zaworra M, Beadle K, Randall E, Hertlein G. et al. Unravelling the molecular determinants of bee sensitivity to neonicotinoid insecticides. Curr Biol. 2018;28:1137-43
154. Heard MS, Baas J, Dorne JL, Lahive E, Robinson AG, Rortais A. et al. Comparative toxicity of pesticides and environmental contaminants in bees: are honey bees a useful proxy for wild bee species? Sci Total Environ. 2017;578:357-65
155. Phan NT, Joshi NK, Rajotte EG, López-Uribe MM, Zhu F, Biddinger DJ. A new ingestion bioassay protocol for assessing pesticide toxicity to the adult Japanese orchard bee (Osmia cornifrons). Sci Rep. 2020;10:9517
156. Phan NT, Joshi NK, Rajotte EG, Zhu F, Peter KA, López-Uribe MM. et al. Systemic pesticides in a solitary bee pollen food store affect larval development and increase pupal mortality. Sci Total Environ. 2024;915:170048
157. Xiao X, Roesner J, Yoon KA, Cho S, Lee SH, Haas J. et al. Unveiling molecular mechanisms mediating coumaphos tolerance in western honey bees. J Agric Food Chem. 2025;73:3389-99
158. Haas J, Glaubitz J, Koenig U, Nauen R. A mechanism-based approach unveils metabolic routes potentially mediating chlorantraniliprole synergism in honey bees, Apis mellifera L, by azole fungicides. Pest Manag Sci. 2022;78:965-73
159. Hayward A, Hunt BJ, Haas J, Bushnell-Crowther E, Troczka BJ, Pym A. et al. A cytochrome P450 insecticide detoxification mechanism is not conserved across the Megachilidae family of bees. Evol Appl. 2024;17:e13625
160. Hayward A, Beadle K, Singh KS, Exeler N, Zaworra M, Almanza MT. et al. The leafcutter bee, Megachile rotundata, is more sensitive to N-cyanoamidine neonicotinoid and butenolide insecticides than other managed bees. Nat Ecol Evol. 2019;3:1521-4
161. Bass C, Hayward A, Troczka BJ, Haas J, Nauen R. The molecular determinants of pesticide sensitivity in bee pollinators. Sci Total Environ. 2024;915:170174
162. Zhu F, Gujar H, Gordon JR, Haynes KF, Potter MF, Palli SR. Bed bugs evolved unique adaptive strategy to resist pyrethroid insecticides. Sci Rep. 2013;3:1456
163. Scott JG, Baker OS, Dressel AE, Norris RH, Burgess ER. Isolation and characterization of the decreased cuticular penetration mechanism of fluralaner resistance in the house fly, Musca domestica. Pestic Biochem Physiol. 2024;205:106154
164. Wang Z, Andika IP, Chung H. Regulation of insect cuticular hydrocarbon biosynthesis. Curr Opin Insect Sci. 2025;67:101287
165. Yan H, Liebig J. Genetic basis of chemical communication in eusocial insects. Genes Dev. 2021;35:470-82
166. Hoffmann K, Gowin J, Hartfelder K, Korb J. The scent of royalty: a P450 gene signals reproductive status in a social insect. Mol Biol Evol. 2014;31:2689-96
167. Sztal T, Chung H, Berger S, Currie PD, Batterham P, Daborn PJ. A cytochrome P450 conserved in insects is involved in cuticle formation. PLoS One. 2012;7:e36544
168. Pridgeon JW, Zhang L, Liu N. Overexpression of CYP4G19 associated with a pyrethroid-resistant strain of the German cockroach, Blattella germanica (L.). Gene. 2003;314:157-63
169. Balabanidou V, Kampouraki A, MacLean M, Blomquist GJ, Tittiger C, Juárez MP. et al. Cytochrome P450 associated with insecticide resistance catalyzes cuticular hydrocarbon production in Anopheles gambiae. Proc Natl Acad Sci U S A. 2016;113:9268-73
170. Chen N, Pei XJ, Li S, Fan YL, Liu TX. Involvement of integument-rich CYP4G19 in hydrocarbon biosynthesis and cuticular penetration resistance in Blattella germanica (L.). Pest Manag Sci. 2020;76:215-26
171. Wang S, Li B, Zhang D. NlCYP4G76 and NlCYP4G115 modulate susceptibility to desiccation and insecticide penetration through affecting cuticular hydrocarbon biosynthesis in Nilaparvata lugens (Hemiptera: Delphacidae). Front Physiol. 2019;10:913
172. Wang YC, Chang YW, Xie HF, Gong WR, Wu CD, Du YZ. The cytochrome P450 gene CYP4g1 driven by high temperature confers abamectin tolerance on Liriomyza trifolii through promoting cuticular hydrocarbons biosynthesis. Pestic Biochem Physiol. 2024;203:106012
173. Kaltenpoth M, Flórez LV, Vigneron A, Dirksen P, Engl T. Origin and function of beneficial bacterial symbioses in insects. Nat Rev Microbiol. 2025;23:551-67
174. Schmidt K, Engel P. Mechanisms underlying gut microbiota-host interactions in insects. J Exp Biol. 2021 224
175. Siddiqui JA, Khan MM, Bamisile BS, Hafeez M, Qasim M, Rasheed MT. et al. Role of insect gut microbiota in pesticide degradation: a review. Front Microbiol. 2022;13:870462
176. Itoh H, Tago K, Hayatsu M, Kikuchi Y. Detoxifying symbiosis: microbe-mediated detoxification of phytotoxins and pesticides in insects. Nat Prod Rep. 2018;35:434-54
177. Fan Q, Sun H, Liang P. The influence of microorganism on insect-related pesticide resistance. Agriculture. 2025;15:1519
178. Zhao M, Lin X, Guo X. The role of insect symbiotic bacteria in metabolizing phytochemicals and agrochemicals. Insects. 2022;13:583
179. Shukla SP, Beran F. Gut microbiota degrades toxic isothiocyanates in a flea beetle pest. Mol Ecol. 2020;29:4692-705
180. Kikuchi Y, Hayatsu M, Hosokawa T, Nagayama A, Tago K, Fukatsu T. Symbiont-mediated insecticide resistance. Proc Natl Acad Sci U S A. 2012;109:8618-22
181. Liu Y, Yu J, Zhu F, Shen Z, Jiang H, Li Z. et al. Function of cytochrome P450s and gut microbiome in biopesticide adaptation of Grapholita molesta on different host diets. Int J Mol Sci. 2023;24:15435
182. Wu Y, Zheng Y, Chen Y, Wang S, Hu F, Zheng H. Honey bee (Apis mellifera) gut microbiota promotes host endogenous detoxification capability via regulation of P450 gene expression in the digestive tract. Microb Biotechnol. 2020;13:1201-12
183. Scates SS, O'Neal ST, Anderson TD. Bacteria-mediated modification of insecticide toxicity in the yellow fever mosquito, Aedes aegypti. Pestic Biochem Physiol. 2019;161:77-85
184. Wang H, Liu H, Peng H, Wang Y, Zhang C, Guo X. et al. A symbiotic gut bacterium enhances Aedes albopictus resistance to insecticide. PLoS Negl Trop Dis. 2022;16:e0010208
185. Cai T, Zhang Y, Liu Y, Deng X, He S, Li J. et al. Wolbachia enhances expression of NlCYP4CE1 in Nilaparvata lugens in response to imidacloprid stress. Insect Sci. 2021;28:355-62
186. Tang T, Zhang Y, Cai T, Deng X, Liu C, Li J. et al. Antibiotics increased host insecticide susceptibility via collapsed bacterial symbionts reducing detoxification metabolism in the brown planthopper, Nilaparvata lugens. J Pest Sci. 2021;94:757-67
187. Pang R, Chen M, Yue L, Xing K, Li T, Kang K. et al. A distinct strain of Arsenophonus symbiont decreases insecticide resistance in its insect host. PLoS Genet. 2018;14:e1007725
188. Sato Y, Jang S, Takeshita K, Itoh H, Koike H, Tago K. et al. Insecticide resistance by a host-symbiont reciprocal detoxification. Nat Commun. 2021;12:6432
189. Selwyn FP, Cheng SL, Bammler TK, Prasad B, Vrana M, Klaassen C. et al. Developmental regulation of drug-processing genes in livers of germ-free mice. Toxicol Sci. 2015;147:84-103
190. Spanogiannopoulos P, Bess EN, Carmody RN, Turnbaugh PJ. The microbial pharmacists within us: a metagenomic view of xenobiotic metabolism. Nat Rev Microbiol. 2016;14:273-87
191. Zemanová N, Lněničková K, Vavrečková M, Anzenbacherová E, Anzenbacher P, Zapletalová I. et al. Gut microbiome affects the metabolism of metronidazole in mice through regulation of hepatic cytochromes P450 expression. PLoS One. 2021;16:e0259643
192. Dempsey JL, Cui JY. Microbiome is a functional modifier of P450 drug metabolism. Curr Pharmacol Rep. 2019;5:481-90
193. Kirsch R, Okamura Y, García-Lozano M, Weiss B, Keller J, Vogel H. et al. Symbiosis and horizontal gene transfer promote herbivory in the megadiverse leaf beetles. Curr Biol. 2025;35:640-54
194. Cheng Z, Chen L, Yan B, Zhang M, Tian J, Mei Y. Insect-eP450DB: A resource of functionally characterized P450s in insects. Insect Sci. 2025;0:1-6
195. Wu H, Li Z, Zhong Z, Guo Y, He L, Xu X. et al. Insect cytochrome P450 database: an integrated resource of genetic diversity, evolution and function. Mol Ecol Resour. 2025;25:e14070
196. Charamis J, Dermauw W, Van Leeuwen T, Vontas J, Feyereisen R. The arthropod P450 enchiridion: An integrated web resource for research on P450s. Insect Biochem Mol Biol. 2025;183:104377
197. Hayward AJ, O'Reilly AO, Nauen R, Bass C, Troczka BJ. An in silico pipeline for enzyme-substrate modelling using arthropod P450s. Pestic Biochem Physiol. 2026;216:106816
198. Kewedar S, Chen Q-R, Moural TW, Lo C, Umbel E, Forrence PJ. et al. Acaricide resistance monitoring and structural insights for precision Tetranychus urticae management. Insects. 2025;16:440
199. Abramson J, Adler J, Dunger J, Evans R, Green T, Pritzel A. et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature. 2024;630:493-500
200. Lomize AL, Todd SC, Pogozheva ID. Spatial arrangement of proteins in planar and curved membranes by PPM 3.0. Protein Sci. 2022;31:209-20
Author contact
![]() Corresponding author: Dr. Fang Zhu, Department of Entomology and Huck Institutes of the Life Sciences, Pennsylvania State University, University Park, PA 16802, USA. Phone: +1-814-863-4432 Fax: +1- 814-865-3048 E-mail: fuz59edu.
Corresponding author: Dr. Fang Zhu, Department of Entomology and Huck Institutes of the Life Sciences, Pennsylvania State University, University Park, PA 16802, USA. Phone: +1-814-863-4432 Fax: +1- 814-865-3048 E-mail: fuz59edu.